Etológia
Kemenes György
tudományos tanácsadó, MRC kutatócsoport vezető, Sussexi Idegtudományi Kutatóközpont, Brighton, Egyesült Királyság
A csigaagy nem bicikli, avagy: elemi idegrendszeri folyamatok és a tanulás evolúciósan konzerválódott sejtes-molekuláris mechanizmusainak vizsgálata puhatestű idegrendszereken*
Az elmúlt mintegy ötven évben a gerinctelen, és elsősorban a puhatestű állatok
idegrendszerein végzett vizsgálatok döntő módon járultak hozzá az idegi
ingerületképzés és -terjedés valamint a tanulás és emléknyom-kialakulás
sejtes-molekuláris folyamatainak megértéséhez. E folyamatok közül több olyannak
bizonyult, amely az evolúció során kevés változáson ment keresztül, és így
egyszerűbb idegrendszerekben való vizsgálatukkal fontos betekintést nyerhetünk
a fejlettebb idegrendszerek elemi és komplex működésének alapvető
mechanizmusaiba.
Sok kitűnő külföldi és magyar idegtudós kollégám az emlős agy közvetlen
vizsgálatával próbálja - rendkívül nagy sikerrel - annak legbonyolultabb
funkcióit föltárni (lásd: Agy és tudat. in Magyar Tudomány. 2001. október). Ha
az állati idegrendszereket autókhoz hasonlítanánk, nyilvánvaló, hogy az emlős
idegrendszer és különösen az emberi agy, mondjuk, egy Rolls Royce-nak felelne
meg, és ha egy ilyen luxusautó összes bonyolult funkcióját meg akarjuk ismerni,
ezt valóban csakis annak közvetlen tanulmányozásával érhetjük el. Azt azonban
talán könnyű belátni, hogy ha csak a Rolls Royce mint robbanómotorral
meghajtott jármű általános működési alapelveiről akarunk képet alkotni, ezt
akár egy Trabant alapos vizsgálatával is megtehetjük (még akkor is, ha a
Trabantot sokan nem is tekintik autónak!). Ennek az az előnye is megvan, hogy
ebben az esetben a luxusautóban meglévő bonyolult, de a működés lényegét nem
érintő komponensek és funkciók (fedélzeti számítógép, légkondicionálás,
minibár, stb.) nem nehezítik meg az alapelvek megértését. Ezzel szemben az is
világos, hogy még a legdrágább és legbonyolultabb kerékpár legalaposabb
vizsgálatával sem jutunk közelebb még a legegyszerűbb autó működésének
megértéséhez sem. Ezt azért fontos hangsúlyoznunk, mert még az élő
természettudományok iránt általában érdeklődő, de az idegtudományokban nem
járatos közönség körében is elterjedt az a nézet, hogy az emlős, és különösen
az emberi agy, az idegrendszer fejlődésének olyan magas szintjét képviseli,
amelyhez képest, hogy az autó hasonlatnál maradjunk, az egyszerűbb, különösen a
gerinctelen, idegrendszerek legföljebb biciklinek tekinthetők, tehát olyan
járműnek, amelynek működését hiába vizsgáljuk, tapodtat sem jutunk közelebb az
autó működésének megértéséhez.
Hogy a fenti nézet helytelen, azt már a zseniális, Nobel-díjas spanyol
neuroanatómus, Santiago Ramón Y Cajal, az ún. neurontan, és így a modern
idegtudományok alapító atyja is felismerte, annak ellenére, hogy a XIX.-XX.
század fordulóján, amikor korszakalkotó kutatásait végezte, a funkció
vizsgálatára közvetlen alkalmas elektrofiziológiai módszerek még nem álltak a
gyakorlatban rendelkezésre (bár az olasz Luigi Galvani már a XVIII. század
végén fölfedezte az állati elektromosságot, és a német Emil du Bois-Reymond,
Johannes Müller és Hermann von Helmholtz már a XIX. században kimutatták, hogy
egy idegsejt elektromos aktivitása meghatározott módon befolyásolja a közelében
lévő többi sejtet). Cajal még a legegyszerűbb féregidegrendszer és a magasan
fejlett emlős agy közötti különbséget is csak olyan bonyolultságbeli
különbségnek tartotta, amely mondjuk egy egyszerű zsebóra és egy csillagászati
óra között van; bár az utóbbi rendkívüli pontossággal mutatja a másodperceket,
perceket és órákat, sőt a Föld pozícióját és mozgását más bolygókhoz képest, az
előbbi is tökéletesen ellátja az órák alapvető feladatát, amely az idő
múlásának mérése. Ugyanígy, érvelt Cajal, a legegyszerűbb gerinctelen
idegrendszer is tökéletesen alkalmazkodott azokhoz a környezeti feltételekhez
amelyek között az adott gerinctelen állatfaj eddigi evolúciója zajlott. Ennek
funkcionális kifejeződése az, hogy még a legegyszerűbb idegrendszerek is
képesek betölteni az idegrendszer legfontosabb általános feladatát - az
organizmust alkotó többi szervrendszer működésének a belső és külső környezet
gyakran változó követelményeinek megfelelő összehangolását.
Egy másik ezzel összefüggő fontos gondolat a darwini szelekciós elméletre
vezethető vissza. A törzsfejlődés során több olyan sejtes-molekuláris
szabályozási mechanizmus alakult ki korán, tehát a gerinctelenek és gerincesek
közös őseinek szintjén, amely azután "kiállta az idő próbáját" (tehát ellenállt
a szelekciós nyomásnak), és emiatt legföljebb csak kis változásokon ment át
mind a gerinctelen, mind a gerinces evolúció során. Jó példa erre az
embrionális testfejlődést megszabó, ún. homeobox génegyüttes, amelyet az
ecetmuslicában (Drosophila sp.) fedeztek fel, és amelyhez felépítésben és
funkcióban nagymértékben hasonló génegyütteseket találtak később az egér és az
ember genomjában is. Már Cajal is felvetette azt a gondolatot, hogy az
idegrendszer törzsfejlődése során is számos ilyen "konzerválódott" mechanizmus
alakult ki, és ezek férgeken, puhatestűeken és rovarokon végzett vizsgálatával
közelebb juthatunk az ilyen mechanizmusok megértéséhez az emlős agyban is.
A jelen tanulmánynak az a célja, hogy immár a Cajal óta fölhalmozódott
hihetetlen nagy mennyiségű új ismeret hátterében bemutassa, hogy a puhatestű
idegrendszerek (amelyeken sokfajta nagy sikerű idegélettani kutatómunka folyik
szerte a világban) és a legfejlettebb emlős idegrendszerek (beleértve az emberi
agy) működési alapelvei azonosak, és hogy ez az azonosság nemcsak a
legalapvetőbb idegrendszeri jelenségekre, hanem a tanulás és memóriakialakulás
bizonyos molekuláris folyamataira is kiterjed. Ennek azért van nagy
jelentősége, mert minden idegrendszer - beleértve az emberi agyat - talán
legbonyolultabb folyamatai éppen a tanulási jelenségekkel függnek össze, és
ezek sejtes/molekuláris alapelveinek feltárása az egyes idegsejtek szintjén még
ma is sokkal könnyebb a számban kisebb, de méretben jelentősen nagyobb
idegsejtekkel rendelkező puhatestűekben, mint a rendkívül bonyolult
idegrendszerrel bíró emlősökben.
A puhatestű állatok óriás idegsejtjeit már a XIX. században fölfedezték, de az
idegélettani vizsgálatok szempontjából igazi jelentőséget csak a múlt század
30-as éveiben nyertek. Miután a 20-as években az angol Lord Edgar Adrian
fölismerte, hogy az idegsejtek elektromos működésének alapegysége az
idegimpulzus vagy akciós potenciál, 1936-ban a szintén angol John Z. Young
fölfedezte a neurofiziológiai kutatások számára a tintahal több mint 1 mm
vastag és 10 cm hosszú óriás axonokkal rendelkező idegsejtjeit (amelyeket
először 1909-ben az amerikai L. W. Williams írt le). Míg Williamsé volt e
sejtek felfedezésének érdeme, Youngnak tulajdonítható az az alapvető
felismerés, hogy az óriás axonok rendkívül alkalmas kísérleti objektumok olyan
kísérletekhez, amelyeknek célja az idegrendszer működésének megértése. Ezen a
nagy jelentőségű de meglehetősen általános felismerésen kívül Young az elsők
között mutatott rá arra is, hogy ha meg akarjuk ismerni az akciós potenciál
keletkezésének mechanizmusát, ezt csak úgy lehet elérni, ha egy elektródával az
axon belsejéből vezetjük el, és így mérjük a sejtmembránon keresztül föllépő
elektromos változásokat.
Young felismerése után nem sokkal ezt a gyakorlatban az amerikai Kenneth Cole
és Howard Curtis, valamint az angol Alan Hodgkin (aki Cole-tól és Curtis-től
"leste el" a tintahal axon használatát, és vezette azt be a brit
laboratóriumokban), Andrew Huxley és Bernard Katz meg is valósította. Nekik
tulajdoníthatók az első intracelluláris elvezetések, amelyeken az akciós
potenciál alakja és a hátterében álló membránkonduktancia-változások egyaránt
tisztán láthatók. Cole fejlesztette ki a feszültségzár (voltage-clamp)
módszert, amelyet aztán Hodkin, Huxley és Katz óriási sikerrel használt a
nyugalmi és akciós potenciál és az ingerületterjedés ionos mechanizmusainak
leírásához.
1952-ben jelent meg Hodkin és Huxley korszakalkotó cikksorozata (az egyik
cikkben Katz is szerzőtárs volt), amelyben részletesen leírták a tintahal axon
használatára épülő kísérleteiket. Az idegsejt elektromos működésének részletes
megértését először lehetővé tevő eredményeikért 1963-ban Hodgkint és Huxleyt
Nobel-díjjal jutalmazták. Az elektrofiziológiai kísérleti technikák további
fejlődésével lehetővé vált, hogy mások gerincesek idegsejtjein is hasonló
méréseket végezzenek el, és ez hamarosan világossá tette, hogy minden idegsejt,
beleértve az emberi agy sejtjeit, lényegében azonos módon tartja fönn a
nyugalmi potenciált, hozza létre és közvetíti az akciós potenciálokat. 1977-ben
Hodgkin (akivel a 80-as évek vége felé a brit Élettani Társaság egyik
előadóülésén még nekem is volt szerencsém találkozni) a következőt jegyezte
meg: "az, hogy Young 1936-ban bevezette a tintahal idegrost kísérleti
használatát, nagyobb mértékben járult hozzá a neurofiziológia fejlődéséhez,
mint bármilyen más felfedezés az elmúlt negyven év során".
A tintahal kísérleti állatként való használata még további két Nobel-díjjal
jutalmazott munka sikeréhez járult hozzá. Az egyik az amerikai George Wald
nevéhez fűződik, akit 1967-ben tüntettek ki a látás kémiai és fiziológiai
folyamatainak összehasonlító élettani szemléletű feltárásáért. Az ő munkássága
azt támasztotta alá, hogy az idegrendszer alapvető működési módjai és azok
kémiai és élettani háttere még az olyan specializált idegsejtek, mint a retina
sejtjei szintjén is nagymértékben konzerválódtak az evolúció során. A másik
Nobel-díjas (1970) az angol Sir Bernard Katz volt (ugyanő vett részt Hodgkinék
eredeti kísérleteiben is), aki a tintahal ideg-izom kapcsolat vizsgálatával
tisztázta a kémiai ingerületátvitel (neurotranszmisszió) alapvetően fontos
kérdéseit, elsősorban a kémiai átvivőanyag-felszabadulás kvantális jellegét. A
japán Hagiwara Szuszumu és az amerikai Theodore H. Bullock a tintahal idegi
szinapszisának vizsgálatával írta le elsőként a kémiai áttevődésű szinapszis
működését. A későbbi vizsgálatok megállapították, hogy a kémiai szinapszisok a
gerinces állatokban is ugyanilyen elevek alapján működnek, és a kémiai
átvivőanyagok nagy részét (például szerotonin, dopamin, acetilkolin,
glutaminsav) mind a gerinctelen, mind a gerinces állatok idegrendszerében
föllelhetjük.
A múlt század 50-es éveinek vége felé már nyilvánvaló volt tehát, hogy az elemi
neuronális mechanizmusok és az ingerületáttevődés terén az egyszerűbb (köztük a
puhatestű) és bonyolultabb idegrendszerek (köztük az emberi agy) is alapvető
azonosságot mutatnak.
Ez a fontos felismerés nyitotta meg az utat olyan új kutatások előtt, amelyek a
puhatestű idegrendszerek működésének elemzésével az elemi jelenségek
vizsgálatának továbbfejlesztése mellett a magatartás idegi
szabályozó-mechanizmusainak feltárására irányultak. Erre a célra különösen a
Gastropodák központi idegrendszere bizonyult alkalmasnak, ahol gyakran
találhatunk óriás, és több fajban (Aplysia, Lymnaea) élénken pigmentált
idegsejteket, amelyeket elektrofiziológiai kísérletekben a francia Angélique
Arvanitaki alkalmazott először. A Gastropoda idegrendszerek vizsgálatán alapuló
neurobiológiai kutatások a múlt század hatvanas éveiben indultak el több
európai laboratóriumban, elsők között a Magyar Tudományos Akadémia Tihanyi
Biológiai Kutatóintézetében (jómagam is itt kezdtem kutatói pályámat, és ahol,
néhány hosszabb-rövidebb külföldi úttal megszakítva 1979 és 1990 között
folytattam kutatásaimat). A hatvanas években létrehozott új laboratóriumokban
(Amszterdam, Jan Lever; Párizs, Ladislav Tauc; Southampton, Gerald Kerkut;
Moszkva, Dimitrij Szaharov; Tihany, Salánki János és S. Rózsa Katalin)
megkezdett puhatestű neurobiológiai munkák korai eredményei is alátámasztották,
hogy számos a gerincesekben is föllelhető elemi és bonyolultabb idegi
szabályozási mechanizmus hatékonyan vizsgálható puhatestű idegrendszerekben.
A tanulás evolúciósan konzerválódott molekuláris mechanizmusainak későbbi
megértését talán legjobban elősegítő munka is igazából e laboratóriumok
egyikéből indult el, bár később főleg amerikai kutatóhelyeken bontakozott ki.
1961-ben egy fiatal amerikai pszichiáter, Eric R. Kandel meglátogatta Tauc
párizsi laboratóriumát, ahol közösen fedezték fel a heteroszinaptikus
facilitáció jelenségét az Aplysia nevű tengeri meztelen csiga faj idegsejtjeit
vizsgálva. Ennek az a lényege, hogy két idegsejt közötti szinaptikus kapcsolat
erőssége megnő, ha előzőleg az egyik (ún. preszinaptikus) sejt szinaptikus
végződése erős bemenetet kap egy harmadik sejt szinaptikus végződésén át.
Ezt követően az Egyesült Államokban a 70-es évek elején óriási lendülettel
indultak meg azok a munkák amelyek Aplysiában a habituáció, szenzitizáció és
klasszikus elkerülési kondicionálás sejtes mechanizmusait először írták le az
ún. "egyszerű rendszer" (simple systems), vagy más néven redukcionista
megközelítési módra építve (Eric R. Kandel, Irving Kupfermann, Vincent
Castellucci, Harold Pinsker, Tom Carew, Jack Byrne, Robert Hawkins, Tom Abrams,
Terry Walters, Craig Bailey, Mary Chen és sokan mások). Ezzel párhuzamosan egy
másik tengeri meztelen csiga fajban (Hermissenda crassicornis) is sikerült
tisztázni egy kondicionált elkerülési válasz neurobiológiai alapjait (Daniel
Alkon, Terry Crow és mások). Ez volt az az időszak, amikor emlősállatokban is
nagy lendületet vettek a tanulási mechanizmusok megértésére irányuló
elektrofiziológiai kísérletek (lásd például Baranyi Attila és Fehér Ottó immár
klasszikusnak számító kísérleteit a macska mozgatókérgének piramissejtjein), és
ezek, a már említett puhatestűeken végzett kísérletekkel együtt a 80-as évek
végére elvezettek ahhoz a felismeréshez, hogy mint az elemi folyamatok terén,
úgy a tanulás sejtes folyamataiban is sok közös vonás van a puhatestű és emlős
idegrendszerek között. A következő általánosnak tűnő alapelvek bontakoztak ki:
1. Az idegrendszeri plaszticitás (tanulásképesség) már meglévő
idegsejthálózatokban lezajló változásokhoz köthető.
2. A plaszticitás előfordulása nem korlátozódik egyetlen idegrendszeri
területre vagy idegsejttípusra.
3. A rövid ideig tartó plasztikus változások ún. második hírvivő rendszerek
működésével és ioncsatornák (főként kálium- és kalciumcsatornák) módosulásával
függnek össze. A második hírvivő rendszerek megnövekedett aktivitása olyan
hosszabb idejű változásokat is triggerelhet, amelyek a tartós emléknyomok
kialakulásában játszanak szerepet (1. ábra).
4. A plasztikus változások nem korlátozhatók egy meghatározott idegrendszeri
területre vagy neuronra, és nem korlátozhatók egyetlen fajta speciális
membránfolyamatra sem. A különböző típusú kifelé irányuló káliumáramok
megváltozása és az ezzel összefüggésben álló, fokozódó kálciumbeáramlás
hatására megnövekedett transzmitter-felszabadulás (1. ábra) azonban olyan
általános mechanizmusnak tűnik, amely inkább csak részleteiben különbözik az
egyes fajok között.
Bár a felsorolt közös vonások a gerinces és gerinctelen idegrendszer tanulási
hátterében álló sejtes mechanizmusokban szembetűnőek, nehezen eldönthető, hogy
ezek is evolúciósan erősen konzerválódott alapvető folyamatokra épülnek-e vagy
csak hasonló funkciójú, de az evolúció során párhuzamosan kialakult jelenségek.
Más a helyzet bizonyos típusú tartós emléknyomok kialakulásának sejtszinten
lezajló folyamatai terén, amelyek minden eddig leírt esetben hasonló típusú
gének aktiválásával, új mRNS- és fehérjeszintézissel járnak. A nyolcvanas évek
közepe óta rohamosan fejlődő molekuláris biológiai módszerek használatával
megint csak az Aplysiában sikerült először feltárni azokat a molekuláris
mechanizmusokat, amelyek tartós emléknyom kialakulásához vezetnek, legalábbis
egyes idegsejtek szintjén (Kandel 2001). Ez jelentősen hozzájárult ahhoz, hogy
Eric R. Kandel munkásságát 2000-ben Élettani és Orvostudományi Nobel-díjjal
jutalmazták, és ez a puhatestű állatok idegrendszerén folytatott vizsgálatok
fontosságának rendkívül magas szintű elismerését jelzi.
A tartós emléknyom kialakulása során a kulcsfontosságú lépések közé tartozik a
ciklikus AMP által közvetlenül vagy közvetve aktivált kináz típusú enzimek
(protein kináz A, MAP kináz) sejtmagba való belépése, ahol azok ún.
transzkripciós (RNS átírást beindító) faktorokat (például CREB, ciklikus AMP-re
válaszoló elemet (CRE) kötő protein) képesek aktiválni (1. ábra). Ezeknek az a
rendkívül fontos szerepük, hogy különböző típusú addig hallgatag gének CRE
szakaszaihoz kötődve azokat aktiválják, és így beindítják azokat a többlépcsős
molekuláris folyamatokat, amelyek új hírvivő RNS és fehérje szintéziséhez és
végső soron a neuronok közötti szinaptikus kapcsolatok tartós megerősödéséhez
vezetnek (1. ábra). Az elmúlt évtized nagy jelentőségű felfedezése volt az,
hogy egyrészt a CREB proteineket kódoló gének felépítése rendkívül hasonló a
csigáktól elkezdve az ecetmuslicán át az egérig és az emberig, másrészt, hogy a
CREB által aktivált gének és további molekuláris mechanizmusok is rendkívül
erősen konzerválódtak (Milner et al., 1998).
Az Aplysiához és Drosophilához hasonlóan a Lymnaea stagnalis nevű csigafaj
(nagy mocsári csiga), amelyen jómagam folytatom az asszociatív tanulás
sejtes-molekuláris megértését célzó vizsgálataimat, CREB proteint kódoló génje
mintegy 85 % homológiát mutat az emlős CREB gén szerkezetével, és ún.
foszforilációs és CRE kötő szakaszai gyakorlatilag azonosak a hasonló funkciót
betöltő emlős CREB génszakaszokkal. Ez azért is érdekes, mert a Lymnaea
táplálkozási magatartását etológiai módszerekkel vizsgálva mi mutattuk ki
először, hogy puhatestű állatok is képesek addig csak gerincesekben megfigyelt
komplex asszociatív tanulásra, amely tartós emléknyom kialakulásával is jár
(Kemenes - Benjamin, 1989). Ez a tanulás a táplálkozási válasz érintési és
kémiai ingerekkel történő kondicionálására épül. Az ilyen ún. appetitív
tanulásnak az az etológiai jelentősége, hogy eredményeképpen a vízicsigák
természetes környezetükben, ahol sokféle növényi anyaggal táplálkoznak, képesek
például már az útjukba kerülő növényi anyag textúrája alapján eldönteni, hogy
az ehető-e vagy nem, és ha igen, milyen tápanyagokban gazdag. Tudományos
szempontból az ilyenfajta tanulási jelenségek vizsgálata egyszerűbb
idegrendszerrel rendelkező állatokban azért rendkívül gyümölcsöző, mert szemben
az Eric R. Kandel által is alkalmazott ún. redukcionista megközelítési móddal,
a magatartási szintről kiindulva, sőt azzal párhuzamosan teszi lehetővé a
tanulás idegi szabályozási folyamatainak vizsgálatát. Az általunk vizsgált
Lymnaeában például a háromfázisú ritmusos táplálkozási mozgásokat létrehozó
idegrendszeri hálózat olyan részleteiben ismert (Staras et al., 1998, 2003;
Straub et al., 2002). hogy ez lehetővé teszi azonosított idegsejtekben lezajló
elektrofiziológiai változások nyomonkövetését a tanulás során (Kemenes et al.
1997; Staras et al., 1999; Benjamin et al., 2000).
A fenti neuroetológiai megközelítési mód alkalmazásával nyilvánvalóvá vált,
hogy nemcsak a gerinctelen állatokon eddig leggyakrabban vizsgált egyszerű
nem-asszociatív (habituáció, szenzitizáció), de bonyolultabb asszociatív
tanulási folyamatok általános molekuláris alapelvei is föltárhatók puhatestű
állatok idegrendszereinek vizsgálatával. Ezen a téren mára már jelentős új
eredményeket értünk el, amelyek először igazolták, hogy a Lymnaea asszociatív
tanulási folyamataiban az emlősökben megfigyeltekhez azonos módon fontos
szerepet játszik a protein kináz A és a MAP kináz, valamint a protein kináz
A-val a CREB aktiválása során szinergista hatású nitrogén oxid (NO) és ciklikus
guanozin monofoszfát (cGMP) (Kemenes et al., 2002).
A korai, főleg Aplysián végzett elemzések szerint az asszociatív tanulás során
lezajló szinaptikus változások ún. nem-hebbi típusúak. 1949-ben Donald Hebb
kanadai pszichológus posztulálta azokat a változásokat, amelyek a szinaptikus
kapcsolatok megerősödéséhez vezetnek. Ennek a folyamatnak lényege az, hogy két
szinaptikusan kapcsolt sejt időben egybeeső aktivitása a szinaptikus kapcsolat
megerősödéséhez vezet. Az ilyen típusú szinapszisok a szakirodalomban hebbi
szinapszis néven váltak ismertté. Terje Lřmo norvég tudós 1966-ban fedezte fel
a tartós potencírozódás (long-term potentiation, LTP) jelenségét az
emlősagyban, amelyet aztán 1973-ban angol kollégájával, Timothy Bliss-szel
együtt írt le részleteiben. Hamar világossá vált, hogy szemben az Aplysiában
leírt ún. aktivitás-függő preszinaptikus facilitációval, amelyről úgy vélik,
hogy ez a klasszikus kondicionálás sejtes mechanizmusa, az LTP hebbi típusú
asszociatív folyamat, amelynek kialakulása során mind a pre- mind a
posztszinaptikus sejt működése rendkívül fontos szerepet játszik. Mivel az LTP
(és LTD, tartós depresszió) eleddig az egyetlen olyan ismert sejtes mechanizmus
az emlősagyban, amely elvileg asszociatív tanulást közvetíthet, úgy tűnt, hogy
mégiscsak lényegi eltérések lehetnek a puhatestű és emlős sejtes asszociatív
tanulási folyamatok között. A kilencvenes években azonban kiderült, hogy
megfelelő kondicionálási paradigma alkalmazásával Aplysiában is fölépíthető
olyan asszociatív tanulás, amely hebbi típusú változásokon alapul, sőt az
emlősagyhoz hasonlóan, a glutaminsav mint pre-szinaptikus transzmitter és
N-methyl-D-aszpartát (NMDA) típusú posztszinaptikus glutaminsav receptorok
működésére épül (Murphy et al., 1997).
A puhatestű állatokban, így az általunk használt Lymnaeában is, az eddig
vizsgált tanulási mechanizmusok mind olyan memóriaformák kialakulásával
függenek össze, amelyek az ún. nem-deklaratív vagy implicit emléknyomok példái.
Ezek olyan tanulási formák eredményeképpen jönnek létre, mint a habituáció,
szenzitizáció, pavlovi és operáns kondicionálás vagy különböző motoros
készségek elsajátítása. Attól messze vagyunk még, hogy az emlős- és különösen
az emberi agyra jellemző rendkívül bonyolult, ún. epizodikus memória
kialakulásának és fennmaradásának molekuláris hátterét is megértsük, és nem
valószínű, hogy ebben a gerinctelen, és köztük a puhatestű modellek közvetlenül
hasznosak lehetnek. Valószínű azonban, hogy a legbonyolultabb memóriafunkciók
is olyan alapkövekből épülnek föl, amelyeket már az evolúció során korán
megjelent, a gerinctelen és gerinces állatokban egyaránt működő tanulási
folyamatok is alkalmaztak, és amelyek jó részét már eddig is puhatestű
idegrendszerek vizsgálatával sikerült feltárni. Ezért remélem, hogy e tanulmány
elolvasása után az olvasó elfogadja, hogy a csigaagy még az emberi agy Rolls
Royce-ához képest sem csak biciklinek tekinthető, és így további vizsgálatával
még több hasznos információt nyerhetünk a legbonyolultabb agyi működések
sejtes-molekuláris alapmechanizmusairól is. Az, hogy az emberi aggyal
összevetve aztán a csigaagy az autók közül is tényleg csak a Trabanthoz
hasonlítható-e, majd remélhetőleg kiderül további kutatásaink során.
Kulcsszavak: Asszociatív tanulás, klasszikus kondicionálás, hebbi szinapszis,
CREB fehérje, Lymnaea stagnalis
Köszönetnyilvánítás
A szerző kutatásait az Orvostudományi Kutatási Tanács (MRC, Egyesült Királyság)
támogatja.
Ezt a tanulmányt szüleimnek, dr. Kemenes Egonnak és dr. Kemenes Egonnénak
valamint feleségemnek, Dr. Kemenesné dr. Kiss Ildikónak ajánlom sok szeretettel.
Irodalom
Benjamin, Paul R. - Staras, Kevin - Kemenes György (2000): A Systems Approach
to the Cellular Analysis of Associative Learning in the Pond Snail Lymnaea.
Learning and Memory. 7, 124-131.
Kandel, Eric R. (2001): The Molecular Biology of Memory Storage: A Dialogue
between Genes and Synapses. Science. 294, 1030-1038.
Kemenes György - Benjamin, Paul R. (1989): Appetitive Learning in Snails Shows
Characteristics of Conditioning in Vertebrates. Brain Res. 489, 163-166.
Kemenes György - Staras, Kevin - Benjamin Paul R. (1997): In Vitro Appetitive
Classical Conditioning of the Feeding Response in the Pond Snail Lymnaea
Stagnalis. Journal of Neurophysiology. 78, 2351-2362.
Kemenes Ildikó - Kemenes Gy. - Andrew, R. J. - Benjamin, P. R. - O'Shea, M.
(2002): Critical Time-Window for NO-Cgmp Dependent Long-Term Memory Formation
after One-Trial Appetitive Conditioning. Journal of Neuroscience. 22, 1414-1425.
Milner, Brenda - Squire, Larry R. - Kandel, Eric R. (1998): Cognitive
Neuroscience and the Study of Memory. Neuron. 20, 445-468.
Murphy, Geoffrey G. - Glanzman, David L. (1997): Mediation of Classical
Conditioning in Aplysia Californica by Long-Term Potentiation of Sensorimotor
Synapses. Science. 278, 467-471.
Staras, Kevin - Kemenes György - Benjamin, Paul R. (1998): Pattern-Generating
Role for Motoneurons in a Rhythmically Active Neuronal Network. Journal of
Neuroscience. 18, 3669-3688.
Staras, Kevin - Kemenes György - Benjamin, Paul R. (1999): Cellular Traces of
Behavioral Classical Conditioning Can Be Recorded at Several Specific Sites in
a Simple Nervous System. Journal of Neuroscience. 19, 347-357.
Staras, Kevin - Kemenes I. - Benjamin, P. R. - Kemenes Gy. (2003): Loss of
Self-Inhibition Is a Cellular Mechanism for Episodic Rhythmic Behavior. Current
Biology. 13, 116-124.
Straub, Volko A. - Staras, K. - Kemenes Gy. - Benjamin, P. R. (2002):
Endogenous and Network Properties of Lymnaea Feeding Central Pattern Generator
Interneurons. Journal of Neurophysiology. 88, 1569-1583
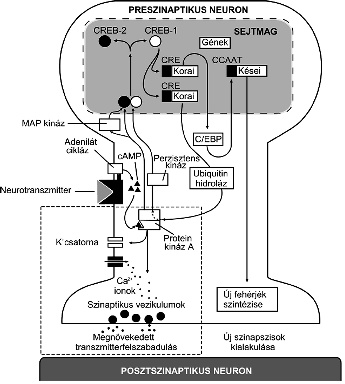 1. ábra * A rövid idejű és tartós szenzitizáció kialakulásához vezető
molekuláris folyamatok preszinaptikus komponenseinek áttekintése az Aplysia
idegrendszer vizsgálatával elért eredmények alapján. Az ábra egy szenzoros
neuront mutat sémás változatban. A rövid idejű szenzitizáció kialakulásának
folyamata (szaggatott vonallal határolt négyszögben) a preszinaptikus végződés
membránjában lévő káliumcsatornák protein kináz A enzim által való átmeneti
foszforilációjára épül. Ezt a szenzitizáló ingerületi pálya neurotranszmittere
váltja ki az adenilát cikláz enzim aktiválásával, amely cAMP képződését
eredményezi. A cAMP a protein kináz A enzim szabályozó alegységéhez kötődve
aktiválja annak katalitikus alegységét, és ezzel alkalmassá teszi azt kálium
ioncsatorna-fehérjék foszforilálására. A káliumcsatorna foszforilációja annak
bezáródásához, így az akciós potenciál időtartamának megnövekedéséhez, és az
így hosszabb ideig nyitva maradó feszültség által aktivált kálciumcsatornákon
át fokozott kálciumbeáramláshoz vezet. Ez megnövekedett
transzmitter-felszabadulást és így erősebb posztszinaptikus választ eredményez,
amelynek a végeredménye fokozott magatartási reakció lesz egy előzőleg gyenge
hatású ingerre. Mindezek a folyamatok az idegsejt sejtmagon kívüli részeiben,
elsősorban a szinaptikus végződésben zajlanak le. A tartós memória kialakulása
ezzel szemben olyan folyamatokhoz kötött, amelyek jórészt a sejtmagban zajlanak
le, és génaktiválással járnak. Ezek a folyamatok a MAP kináz és a protein kináz
A enzimek sejtmagba való belépésével kezdődnek. A sejtmagba belépve ezek az
enzimek aktiválják a CREB ún. aktivátor változatát (CREB-1) míg inaktiválják az
ún. represszor változatot (CREB-2). Az aktivált CREB korai gének CRE
szakaszához kötődve azok működését beindítja, és ez a C\EBP nevű újabb
transzkripciós faktoron át kései gének aktiválásához vezet, illetve más korai
gének (pl. ubiquitin hidroláz) aktiválásán át azt eredményezi, hogy a protein
kináz A enzim katalitikus alegysége perzisztensen aktívvá válik. Mindezen
folyamatok végeredménye a megnövekedett fehérjeszintézis, amely új szinapszisok
kialakulásában játszik szerepet. Mind a rövid mind a tartós memória itt
bemutatott molekuláris lépései igen hasonlónak bizonyultak a gerinctelen és
gerinces állatok eddig leírt sejtszintű tanulási modelljeiben.
1. ábra * A rövid idejű és tartós szenzitizáció kialakulásához vezető
molekuláris folyamatok preszinaptikus komponenseinek áttekintése az Aplysia
idegrendszer vizsgálatával elért eredmények alapján. Az ábra egy szenzoros
neuront mutat sémás változatban. A rövid idejű szenzitizáció kialakulásának
folyamata (szaggatott vonallal határolt négyszögben) a preszinaptikus végződés
membránjában lévő káliumcsatornák protein kináz A enzim által való átmeneti
foszforilációjára épül. Ezt a szenzitizáló ingerületi pálya neurotranszmittere
váltja ki az adenilát cikláz enzim aktiválásával, amely cAMP képződését
eredményezi. A cAMP a protein kináz A enzim szabályozó alegységéhez kötődve
aktiválja annak katalitikus alegységét, és ezzel alkalmassá teszi azt kálium
ioncsatorna-fehérjék foszforilálására. A káliumcsatorna foszforilációja annak
bezáródásához, így az akciós potenciál időtartamának megnövekedéséhez, és az
így hosszabb ideig nyitva maradó feszültség által aktivált kálciumcsatornákon
át fokozott kálciumbeáramláshoz vezet. Ez megnövekedett
transzmitter-felszabadulást és így erősebb posztszinaptikus választ eredményez,
amelynek a végeredménye fokozott magatartási reakció lesz egy előzőleg gyenge
hatású ingerre. Mindezek a folyamatok az idegsejt sejtmagon kívüli részeiben,
elsősorban a szinaptikus végződésben zajlanak le. A tartós memória kialakulása
ezzel szemben olyan folyamatokhoz kötött, amelyek jórészt a sejtmagban zajlanak
le, és génaktiválással járnak. Ezek a folyamatok a MAP kináz és a protein kináz
A enzimek sejtmagba való belépésével kezdődnek. A sejtmagba belépve ezek az
enzimek aktiválják a CREB ún. aktivátor változatát (CREB-1) míg inaktiválják az
ún. represszor változatot (CREB-2). Az aktivált CREB korai gének CRE
szakaszához kötődve azok működését beindítja, és ez a C\EBP nevű újabb
transzkripciós faktoron át kései gének aktiválásához vezet, illetve más korai
gének (pl. ubiquitin hidroláz) aktiválásán át azt eredményezi, hogy a protein
kináz A enzim katalitikus alegysége perzisztensen aktívvá válik. Mindezen
folyamatok végeredménye a megnövekedett fehérjeszintézis, amely új szinapszisok
kialakulásában játszik szerepet. Mind a rövid mind a tartós memória itt
bemutatott molekuláris lépései igen hasonlónak bizonyultak a gerinctelen és
gerinces állatok eddig leírt sejtszintű tanulási modelljeiben.
<-- Vissza a 2003/8 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]