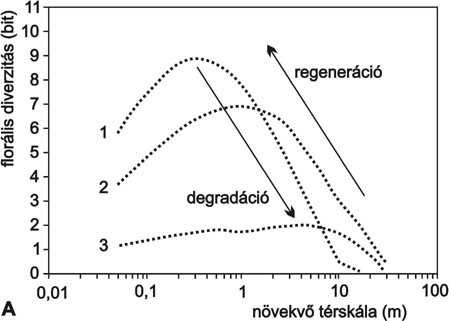
 1. ábra * A koegzisztenciális mintázattranszformációk alapfolyamatai. A -
regeneráció és degradáció; B - egy hazai növénytársulás (Festucetum vaginatae)
koordináltságának csökkenése a globális ökológiai változásokat reprezentáló
gradiens mentén.
1. ábra * A koegzisztenciális mintázattranszformációk alapfolyamatai. A -
regeneráció és degradáció; B - egy hazai növénytársulás (Festucetum vaginatae)
koordináltságának csökkenése a globális ökológiai változásokat reprezentáló
gradiens mentén.
Bevezetés
A növénytakaró a szupraindividuális szerkezetek látszólag jól felismerhető komponense. A vegetációval borított tájban - legalábbis felületes módon - a laikus is jól tájékozódik. Talán erre vezethetők vissza azok az előítéletek, amelyek a vegetáció vizsgálatát könnyű foglalatosságnak, afféle játéknak tartják, és a tudományterületet, mint egyfajta haszontalanságot, igazságtalanul az ún. soft science-ek közé sorolják.
Ezzel szemben történeti tény, hogy a vegetációtan, a növényrendszertannal együtt már a gyarmatosítás korában igen fontos gyakorlati szerepet töltött be egy-egy terület ökológiai potenciáljának és gazdasági hasznosíthatóságának felmérésében. A gyakorlati kihívások második hulláma, az ipari szemléletű mezőgazdaság térnyerésével párhuzamosan, a XIX. sz.második felében jelentkezett, amikor például Svájc, Oroszország és az Amerikai Egyesült Államok legelőinek a túlhasználata óriási területek elgyomosodását, degradációját, s ezzel a tej- és húsprodukció drasztikus visszaesését eredményezte.
Talán nincs még egy diszciplína Magyarországon, amelynek fejlődéséről, eredményeiről ennyire keveset tud a hazai tudományos és közvélemény. Pedig a közelmúltban éppen a vegetációtan területén belül születtek meg azok a felfedezések, amelyek a diverzitás keletkezésének, fennmaradásának és funkciójának teljesen új megközelítéseihez vezettek, felfedték a természetes zavarások és az egyensúlyoktól távoli folyamatok szerepét a természetes vegetáció működésében, és ezzel együtt gyökeresen átértékelték az ember és a természet viszonyáról korábban kialakult nézeteinket. Ezeket az új eredményeket döntően a kísérletes vegetációdinamika és a térben explicit szimulációs modellezés produkálta. Azonban a kialakuló új paradigma a vegetációtan valamennyi részterületét átalakította. Az új szemlélet következtében erősödött az egyes ökológiai szub- és részdiszciplínák közötti kommunikáció és általában a szupraindividuális tudományterületek integrációja.
A legtöbb ember a vegetációt, így például a lakóhelyét körülvevő erdőket és réteket mint a környezetének stabil alkotórészeit érzékeli, sőt, ezekhez képest tájékozódik, mozog a tájban. A növényzet fogalmához ősidők óta a mozdulatlanság, stabilitás és ezzel együtt egyfajta biztonságérzet is társul. Mindannyian azzal a tudattal térünk aludni, hogy a környezetünkben élő fákat, bokrokat holnap reggel is ugyanott találjuk majd. Az utolsó másfél évtized egyik legfontosabb felfedezése, hogy ez a mozdulatlanság csak látszat, a humán percepció sajátságainak a következménye. A vegetáció szempontjából releváns biológiai léptékben, az egyes növényegyedek, klónok illetve populációk, növekedésük, pusztulásuk és utódjaik terjedése következtében állandóan változtatják a helyüket. Ha egy felgyorsított filmen követnénk a növényzet képét, sebesen kavargó, örökösen átformálódó mintázatokat látnánk. E nélkül a dinamikus, hosszú időtartamot (évtizedeket, évszázadokat) átfogó szemlélet nélkül a vegetáció működése, viselkedése, funkciói nem értelmezhetők. A felismerés, hogy a vegetáció működésének léptéke a humán megismerés léptékétől ilyen sajátságosan különbözik, forradalmasította a vegetációtudományt. De a fontos metodológiai konzekvenciákon túl filozófiai következményekkel is jár, és arra kényszerít minket, hogy általában is átértékeljük az élet- és természettudományok viszonyát a Természethez. A biológia tudománya sokáig az élet egységét kereste. A legutóbbi ökológiai felfedezések nyomán azonban rá kellett döbbennünk a sokféleség életünkben játszott meghatározó szerepére. Kiderült, hogy az élővilág korábban csak romantikus kuriózumként, érdekes látványosságként kezelt diverzitása és komplexitása döntő szerepet játszik életfeltételeink kialakításában, meghatározza világunk stabilitását, kiszámíthatóságát és az olyan alapvető értékek hozzáférhetőségét, mint amilyen az elviselhető klíma, a belélegezhető levegő és az iható víz.
Az új paradigma kialakulása
Harminc-negyven évvel ezelőtt a vegetációtanban a leíró, összehasonlító és rendszerező szubdiszciplínák (szünmorfológia, szüntaxonómia) domináltak. A növénytársulást zárt egyensúlyi rendszernek tekintették, amelynek dinamikája döntően a belső mechanizmusok működésén alapul. A kor szemléletét kifejező ún. egyensúlyi paradigma szerint a társulások állapotát a lokális termőhelyi paraméterek (talaj és klíma) és a fajok között a limitált forrásokért folyó versengés határozzák meg. Más trofikus szintek, például a növényevők hatása elhanyagolható. Az ember (illetve a tájhasználat) szerepe csak kivételesen, mint zavaró hatás jelentkezik. Elhanyagolható a múlt és a szomszédos vegetáció. A társulás mindenkori állapotát a fajok átlagos mennyiségeit megjelenítő ún. faj/abundancia textúra jellemzi. A társulást alkotó populációk dinamikája differenciálegyenletekkel modellezhető. A modellek segítségével megjósolható adott vegetációtípus (szüntaxon) ideális (egyensúlyi) összetétele. A fajok együttélésének feltételei az interakciókat jellemző paraméterek (intraspecifikus és interspecifikus kompetíciós koefficiensek) közötti relációkkal adhatók meg.
Ezután egy redukcionista, kísérletező periódus következett, amely kialakította a konkrét hipotéziseket tesztelő, oknyomozó funkcionális és dinamikus szemléletű szubdiszciplínákat (vegetációdinamika, funkcionális vegetációtan, szünfiziológia). A 70-es években kezdődő kísérletes, mechanisztikus irányzat igazi sikertörténetnek indult. Adva voltak a korábbi leíró-összehasonlító korszak által tett megfigyelések és általánosítások, azaz a megmagyarázandó jelenségek, adva voltak a társulás szerkezetét és dinamikáját leíró modellek, és ehhez hamar kialakult a kísérleti módszertan és a szükséges statisztikai eljárások. A megfigyelések, a kísérletek és a modellezés összekapcsolása azonban komoly, nem várt nehézségekbe ütközött. A kísérletes megközelítés keretében tereplaboratóriumokat is kiépítettek, ahol állandósított mintavételi egységekben követték a vegetáció változásait. Mindez pontosabb méréseket eredményezett, amelyek megkérdőjelezték a vegetáció mintázataira és folyamataira korábban megállapított trendeket, törvényszerűségeket. A korábbi becslések a térbeliségből az időbeliségre következtető spekulatív módszerrel történtek. Mivel a releváns vegetációdinamikai folyamatok igen hosszú időtartamúak (évtizedes, évszázados léptékben zajlanak le), ezért a kutatók az egymás szomszédságában fekvő különböző korú területek állapotainak önkényes összeillesztésével rekonstruálták a megfelelő folyamatokat. Az új közvetlen mérések azonban azt mutatták, hogy a rekonstrukciós módszer jelentősen alulbecsülte a folyamatok komplexitását és variabilitását. De hasonló problémák jelentkeztek a mechanizmusok feltárása során is.
A redukcionista megközelítés módszerelméleti posztulátuma, hogy a természetben előforduló komplex és változatos mintázatok egyszerű generatív szabályokra vezethetők vissza. Ennek kapcsán feltételezzük, hogy a populációk számunkra fontos tulajdonságai (például a szaporodó- és versenyképességet a talajnedvesség függvényében leíró összefüggés paraméterei) viszonylag állandók, és ezek a paraméterek kísérletesen meghatározhatók. Ha ismerjük az alkotó populációk tulajdonságait (azaz fajonként kb. tíz-húsz paraméter értékét), akkor bármely fajkombinációra és bármely környezeti feltétel esetére (sok százezer, de akár sok milliárd esetre is) megjósolható a közösség viselkedése. Kezdetben úgy tűnt, hogy a kontrollált üvegházi körülmények között végzett rövid időtartamú mérések igazolják ezt a redukcionista programot (Harper, 1977). Ezeket a méréseket egyetlen populáción vagy egy kisebb, két-három populációból álló mesterséges közösségen végezték, és eredményül egyszerű függvényekkel leírható szabályos viselkedéseket kaptak. Kiderült azonban, hogy az in vitro (üvegházi körülmények között) megállapított populációs paraméterek segítségével a terepen észlelt jelenségek csak ritkán értelmezhetők. Például, ha két növényfajt különböző arányokban elvetünk, és követjük a keverékek populációinak a relatív növekedését, akkor meghatározhatjuk a fajpárra jellemző interspecifikus kompetíciós paramétert. Sokfajú természetes rendszerekben terepi körülmények között végzett mérések azonban azt mutatták, hogy a két faj egymással szembeni viselkedése más fajok jelenlétében megváltozik, tehát a kétfajú kombinációkra kapott eredmények a sokfajú közösségekre nem általánosíthatók.
A kísérletes módszer lényegénél fogva egyszerűsítésre kényszerül. A problémát az okozza, hogy gyakran nem ismerjük az egyszerűsítések következményeit. A kényszerűen rövid időtartamú kísérletek arra nem adhatnak választ, hogy az egyszerűsített körülmények között, in vitro mért tulajdonságok és jelenségek mennyire relevánsak a populációk tartós együttélése szempontjából. Az egyszerűsítő feltételek következményeinek szisztematikus vizsgálata a 80-as évek elejétől, a nagyteljesítményű számítógépek megjelenésével vált lehetségessé. Olyan új, térben explicit, egyedalapú szimulációs modellek születtek, amelyekkel a korábbiaknál sokkal realisztikusabban lehetett megjeleníteni a növényközösségek kompetíciós dinamikáit (Czárán, 1998). A korábbi klasszikus modellekben (az üvegházi kísérletekhez hasonlóan) feltételeztük, hogy az azonos populációhoz tartozó egyedek egyformák, a populációk tökéletesen keverednek, az interakciók pedig nem függnek a populációk méretétől, illetve a közösség számára rendelkezésre álló terület méretétől. Az új típusú modellekkel sikerült bebizonyítani, hogy ezek az egyszerűsítő feltételek egyáltalán nem triviálisak, és megváltoztatásuk döntően megváltoztatja a közösség kompetitív dinamikáját. Ezek az eredmények azért is érdekesek, mert választ adtak az ökológia egyik fontos paradoxonjára. Közel 40 éve vetette fel G. Evelyn Hutchinson azt a problémát, hogy a tavakban együtt élő sok száz algafaj együttélésének ténye ellentmondásban van a niche-elmélet azon állításával, hogy a fajok száma nem haladhatja meg azoknak a forrásoknak a számát, amelyekért versengenek. A vizsgálatok szerint ugyanis a fajok száma több mint egy nagyságrenddel felülmúlta a limitáló tényezők (például bizonyos vízben oldott és tápanyagként hasznosított szervetlen sók) számát. A paradoxon később vizsgálatok ezreit motiválta, és erősen közrejátszott abban, hogy kialakult a nem-egyensúlyi paradigma. Kiderült, hogy a paradoxon oka nem a természetben, hanem a modellekben keresendő. A paradoxon csak akkor áll elő, ha a társulásokat a klasszikus kémiai reakciók mintájára képzeljük el, ahol nagyszámú részecske, híg oldatban, jól összekeveredve, nagy sebességgel reagál, és a reakció egyensúlyra vezet. Ezzel szemben, ha a figyelembe vesszük, hogy az ökológiai reakciók relatíve lassúak, az ökológiai rendszer térfogata véges, és kis egyedszámok is előfordulnak, hogy az egyedek nem egyforma méretűek, illetve hogy a populációk nem keverednek tökéletesen, minden esetben javulnak a koegzisztencia-feltételek az eredeti egyensúlyi modellhez képest.
Az egyensúlyi paradigma feltételezi, hogy a természetes közösségek többsége egyensúlyi vagy ahhoz közeli állapotban van, illetve hogy egy perturbációt követően az egyensúlyok hamar beállnak. Ezzel szemben egyre több esettanulmány kérdőjelezte meg az egyensúlyok létét a természetben, és demonstrálta, hogy a növénytársulásokra többféle (tíz-, száz-, ezeréves) időskálán egyaránt a folyamatos változás, átalakulás a jellemző. Más vizsgálatok azt bizonyították, hogy a társulások zárt rendszerként való felfogása is tarthatatlan. Eredményeik szerint a lokális vegetációdinamikák sokféleségének egyik fontos oka a társuláson kívül keresendő, mégpedig a környező növényzet propagulumforrásainak a jellemzőiben. Kimutatták, hogy bizonyos esetekben a propagulumforrás távolsága jobb prediktora a vegetáció állapotának, mint a terület dinamikai állapota (szukcessziós kora). A legújabb elméletek szerint a környező táj hatása a társulás koegzisztenciális mintázataira és dinamikáira az állományon belüli erőkkel egyenrangú lehet, sőt, azokat esetenként felülmúlja.
A táji léptékű vegetációdinamikai kényszerek szerepének a felismerése mellett az egyensúlyi paradigmának ellentmondott az a megfigyelés is, hogy a védett területek állapota gyakran tovább romlott a szigorú védelem, a további zavarások teljes kizárása ellenére is. Kiderült, hogy a védelem által funkcionálisan valóban zárt rendszerré alakított terület gyakran nem tud a normális módon működni és fennmaradni. Felismerték továbbá, hogy a vegetációt érő természetes zavarások (például a növényevők, a tűz, a viharok) részét képezik a természetes rendszernek, és az általuk okozott vegetációs dinamika szükséges a társulás diverzitásának a fennmaradásához. A diverzitás fennmaradása szempontjából fontosnak bizonyultak a növényi populációk állományon belüli mintázatai és e mintázatok átrendeződései is. Ha a társulás belsejében kijelölünk egy kisméretű mintavételi egységet, azon belül soha nincs egyensúly, hanem az egyedek, és ezzel a fajok folyamatos cseréje figyelhető meg. Bizonyos mintázatdinamikai feltételek teljesülése esetén azonban a lokális fajcserék hatásai egymást kiegyenlítik, és nagyobb léptékben, az állomány szintjén már viszonylag állandó összetételt és a fajok együttélését tapasztaljuk.
Az új eredményekből születő szemlélet, amely az ún. nem-egyensúlyi paradigma néven nyert megfogalmazást (Pickett et al., 1992), az állapotok helyett a folyamatok vizsgálatára helyezte a hangsúlyt. A nem-egyensúlyi paradigma szerint a vegetáció és általában az ökológiai rendszerek nyílt, egyensúlytól távoli rendszernek tekintendők, ahol a lokális folyamatok nem függetlenek egymástól, azaz egy adott objektum viselkedését (időbeli állapotváltozásait) a szomszédos objektumok és azok változásai nagy mértékben befolyásolják. A vizsgált rendszer viselkedése függ saját belső szerkezetétől, és a környezetével való kapcsolatai is mintázatok interakcióiként értelmezhetők. Az új szemlélet az ökológiai kontextusokra, azaz a térbeli és időbeli szomszédsági relációkra, valamint az anyag- és energiaforgalmi (például a táplálékláncbeli) kapcsolatokra helyezte a hangsúlyt. Annak eldöntése, hogy egy adott vizsgálati objektum mint ökológiai alrendszer egyensúlyban van-e, illetve hogy milyen távol van az egyensúlytól, nem is olyan egyszerű kérdés. A válasz függ a vizsgálat térbeli és időbeli léptékétől, az alkalmazott perturbációtól, és attól is, hogy hol húzzuk meg a mesterségesen elkülönített alrendszer határait. Például az észak-amerikai hosszú füvű préri tipikusan nem-egyensúlyi rendszer, mert magára hagyva, zavarás hiányában beerdősül az adott klíma mellett. Ha azonban a nagytestű legelő állatokat és a spontán préritüzeket is a rendszer részének tekintjük (ami evolúciós szempontból indokolt lehet), akkor ugyanez a rendszer egy bizonyos lépték felett egyensúlyinak is tekinthető. Általában igaz, hogy a nagy területű, erős emberi behatásoktól mentes természetes rendszerek (például a boreális erdők) egy bizonyos tér- és időléptékben (időben néhány évtizedes, térben több négyzetkilométeres léptékben) perzisztensnek mondhatók, és kisebb zavarások után az eredetihez közeli állapotba regenerálódnak. Az állományléptékű állandóság azonban a legtöbb esetben élénk, finomabb léptékű dinamikák segítségével valósul meg. E problémák miatt az új paradigma "nem-egyensúlyi" elnevezése nem szerencsés, és talán helyesebb lett volna "mintázatdinamikai paradigmaként" meghatározni. A lényeget azonban nem a tudománytörténeti esetlegességként kialakult címke, hanem a szemléletváltást követő metodológiai változások (1. táblázat) jelentik.
Metodológiai következmények
A korábbi paradigma esetében az egyensúlyi állapotok kitüntetése módszertanilag igen kényelmes egyszerűsítő feltevés volt, hiszen kimondható, hogy egyensúlyban a vegetáció csak az állomány belső, pillanatnyi állapotától és a szorosan hozzá tartozó lokális környezettől függ. Ebben az esetben a modellezés során a térbeli relációk elhanyagolhatók, a koegzisztenciális dinamikát jellemző törvényszerűségek viszonylag egyszerűen, a modell paraméterei közötti egyenlőtlenségekkel megfogalmazhatók. Ezzel szemben a nem-egyensúlyi paradigma esetében a vizsgált jelenségeket téridő mintázatok, illetve téridő folyamatok formájában írjuk le, és ennek megfelelően bonyolultabb reprezentációkat kell alkalmaznunk. Az új szemlélet a statisztikai, mintavételi és dinamikai reprezentációk teljes átalakítását kívánta (1. táblázat).
A vegetáció sok más biológiai rendszerhez hasonlóan hierarchikusan szerveződik. Egyensúly esetén, amikor a szerveződési hierarchia stabil, és az egyes szinteket nagyságrendnyi különbségek választják el, egy konkrét vizsgálatban a kiválasztott objektum mérete és a feltett kérdések automatikusan kijelölik a vizsgálat léptékét. Egyensúlytól távoli rendszerekben azonban a vizsgálati skála kijelölése kevésbé triviális. Időben változó mintázatok esetén a szerveződési hierarchia sajátságai is folyamatosan változnak. Egy adott szint viselkedése részben a magasabb szintű mintázatoktól függ, amelyekbe beágyazódik, részben a komponenseiként megjelenő alsóbb szintű mintázatoktól, amelyek saját belső variációs és ezzel válaszadási képességét meghatározzák. Az egyes részmintázatok képeződése, lebomlása során maga a mintázati hierachia is folyamatosan átalakul. Egyes szintek megjelenhetnek, eltűnhetnek, a szintek közötti távolság változhat, különleges esetben skála-invariáns szerkezetek, fraktálok képződnek. Ebből adódóan nem dolgozhatunk egyetlen, önkényesen választott léptékben. A vizsgálatok során a mintavételi egységek méretét változtatva, ún. térsorozati vagy idősorozati skálázást kell alkalmaznunk, ahol az adott vegetációs attribútum léptékfüggése maga is állapotjelző karakterisztikum.
Említettük, hogy a vegetációtan más szupraindividuális diszciplínákhoz hasonlóan az élő természet sokféleségének a leírásával és értelmezésével foglalkozik. A sokféleség megközelítésének fontos eleme az osztályozás és rendszerezés. Ugyanakkor az osztályozás során fennáll a veszély, hogy az azonos osztályba sorolt objektumok közötti változatosságot elhanyagolva a sokféleséget jelentősen alulbecsüljük. Az egyensúlyi paradigma alapot jelentett ezekhez az egyszerűsítésekhez. Ha a klasszifikáció során elkülönített szüntaxonokat egyensúlyi állapotoknak tekintjük, akkor az ezektől való eltérések, illetve az egyes típusok közötti átmenetek elhanyagolhatók. Feltételezzük ugyanis, hogy a nem-egyensúlyi, átmeneti állapotokat csupán valamilyen időleges perturbáció hozta létre, és ezek a zavaró hatás elmúltával visszaalakulnak majd a megfelelő egyensúlyi állapotba. Az új nem-egyensúlyi paradigma esetében, azáltal, hogy folyamatokat vizsgálunk, és azáltal, hogy a folyamatokat mintázatfüggőnek tekintjük, az átmeneti állapotok szerepe, jelentősége megnő. Ezért olyan állapotreprezentációkat kellett kifejleszteni, amelyek képesek a nagyszámú átmeneti állapot kezelésére. A feladat nagyon nehéz. Addig, míg egy tipikus egyensúlyi esetben húsz-ötven állapot elkülönítése rendszerint elegendő, a megfelelő nem-egyensúlyi esetben az átmenetekkel együtt már sokezer állapot, transzformáció és egyéb reláció megkülönböztetésére van szükség. További komplikációt okoz, hogy az átmenetek száma függ a vizsgálat léptékétől.
A fentiek ismeretében érthető, hogy miért dominálták a vegetációtan elmúlt húsz évét a módszertani vizsgálatok. Szerencsés módon erre az időszakra esik a számítógépes kapacitások jelentős növekedése, a térinformatika, a távérzékelés és az adatbázisok fejlődése is. Bár ezek a technikák jelenleg még messze nem elégítik ki a biológusok igényeit, alkalmazásuk máris forradalmasította a vegetációtant, és lehetővé tette a mintázatokkal való operatív munkát. A vegetáció új szemlélete és az új módszerek hatására megkezdődött a korábban jórészt izoláltan fejlődő szüntaxonómia, szünmorfológia és szündinamika szintézise egy, az eddiginél jóval hatékonyabb, dinamikus és funkcionális vegetációtan keretében.
A módszertani, módszerelméleti fejlesztések közül kiemelkedik Juhász-Nagy Pál munkássága. Neki sikerült a világon először (ráadásul korát harminc évvel megelőzve) összekapcsolni a sokváltozós és a térbeli statisztikai eljárásokat egyetlen koherens modellcsalád formájában. A biológiai komplexitás reprezentálására alkalmas információelméleti modellek kidolgozása mellett megreformálta a klasszikus vegetációtan fogalomrendszerét és módszerelméletét is (Juhász-Nagy, 1980), amely így alkalmassá vált az új paradigma szerinti munkára. A sok ezer állapot és reláció azáltal válik kezelhetővé, hogy a statisztikus mechanikához és a termodinamikához hasonlóan a rendszer átlagos rendezettségi állapotait jellemezzük.
Az 1. ábra két alapvető cönológiai törvényszerűséget mutat be Juhász-Nagy módszereit felhasználva. A florális diverzitás nevű cönológiai állapotváltozó a fajok együttélési módjainak sokféleségét méri az állományon belül. Ha az együttélést semmi nem akadályozza, és a fajok lokális előfordulásaik során szabadon kombinálódnak, akkor a függvény értéke maximális, maximuma pedig kis térléptéknél jelenik meg. Ha az együttélés korlátozott, akkor bizonyos fajkombinációk tiltottak, és a függvény értéke kisebb lesz. Ha egy növényközösséget zavarás ér, akkor először a finom térléptékű együttélések szerkezete bomlik fel, a vegetáció mozaikossá válik. A mozaik egy-egy foltján belül pedig jelentősen lecsökken az együtt előforduló fajok kombinációinak változatossága. Ilyenkor a cönológiai állapotjelző függvény maximuma a nagyobb térléptékek felé tolódik, jelezve, hogy egymáshoz közel, kis térrészletben már nem vagy csak kevésbé képesek együtt élni a fajok. Egy koegzisztenciális szerkezet felbomlása, degradációja során tehát a fajkombinációk diverzitásának csökkenése és a maximális diverzitáshoz tartozó karakterisztikus skála nagyobb térléptékek felé való eltolódása következik be. Egy szukcessziós folyamatban, amikor a társulás regenerálódik, a diverzitás növekedése és a karakterisztikus maximum skála csökkenése, tehát a degradációval ellentétes irányú transzformációk figyelhetők meg (1.A ábra).
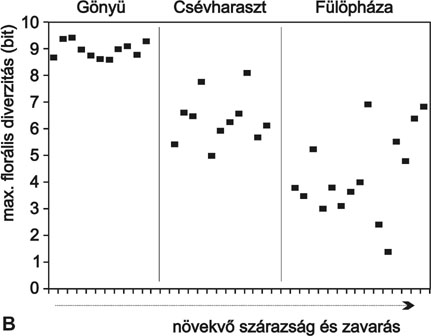
Az 1.B ábra több tucat vegetációfolt állapotát mutatja egy ökológiai gradiens három pontján. (Az áttekinthetőség végett a cönológiai állapotjelző függvényeknek itt csak a maximumait ábrázoltuk, tehát minden pont egy adott vegetációs állomány koegzisztenciális állapotát jellemzi). Itt is látható, hogy az ökológiai gradiens mentén a növekvő emberi zavarás és a növekvő ariditás hatására csökken a florális diverzitás, elszegényedik, leromlik a közösség. Ugyanakkor az is látszik, hogy a zavartabb és szárazabb környezetben a pontok szórása nagyobb, csökken a társulások koordináltsága (homeosztázisa), ami közvetve jelzi a szabályozási, önfenntartó és önreprodukáló mechanizmusok gyengülését. Valamennyi itt felhasznált adat ugyanabból a társulásból, a magyar csenkesz és a pusztai árvalányhaj gyepjeinek szép, értékes, védett állományaiból származik. Ezek finom változásai, a kezdődő problémák a felületes szemrevételezéskor még láthatatlanok, de az általunk használt érzékeny módszer egyértelműen jelzi a riasztó tendenciát. Sajnos a példában bemutatott jelenség általános. A koordináltság csökkenése a jelenlegi globális ökológiai változásokat (növekvő emberi zavarások, szárazodás és egyre gyakoribb időjárási szélsőségek) kísérő törvényszerű cönológiai jelenség.
Cönológiai mintázatok és folyamatok dinamikai összefüggéseinek megértése érdekében érdemes összekapcsolni a Juhász-Nagy-féle sokváltozós térbeli statisztikai eszközöket a térben explicit modellezéssel. Térben explicit egyedalapú szimulációs modellek segítségével absztrakt referenciaközösségek dinamikája modellezhető. Ilyenkor szisztematikusan változtatjuk a fajok dinamikai paramétereit (demográfiai, inváziós, diszperzál, kompetíciós, a zavarásokkal és a környezeti feltételekkel szembeni toleranciát leíró paramétereket), a szimulált mintázatképződési folyamatokat pedig a Juhász-Nagy modelljeivel definiált állapottérben ábrázoljuk. Kimutatható, hogy összefüggés van a generatív paraméterek osztályai és a szimulált trajektórium halmazai között (Bartha et al., 1998). A cönológiai állapottérnek a fenti módszerekkel való beskálázása lehetővé teszi, hogy távoli területekről származó egyedi terepi mérések adatait összehasonlítsuk, és mintázatdinamikai, társulásszerveződési kontextusban értékeljünk (2. ábra).
A szimulációs eredményeket a terepadatokkal összevetve jó egyezést tapasztaltunk a szimuláció által jósolt mintázatképző mechanizmusok és az adott terepi objektumról a szakirodalomban leírt releváns mechanizmusok között (Kovács-Láng et al., 1999; Bartha, 2000).
Funkcionális vegetációtan
A funkcionális vegetációtan azokkal a folyamatokkal foglalkozik, amelyek révén a növényzet anyagokat, energiát (és információt) köt meg, alakít át, tárol vagy transzportál. E folyamatok következménye a környezethez való adaptáció, a zavarások utáni regeneráció, az önfenntartás és önreprodukció. Ennek során a növényzet nemcsak alkalmazkodik környezetéhez, de azt sok esetben átalakítja, egyben hozzájárul az ökoszisztéma illetve a bioszféra működéséhez (Tuba et al., 1998).
A különböző környezettípusokban megjelenő változatos növényi adaptációs formák kérdése régóta foglalkoztatja a kutatókat. Az új kutatási paradigma fontos következménye, hogy az utóbbi években az érdeklődés egy adott környezeti típuson belül is az átlagok helyett a variáció és a mintázatok leírása és értelmezése felé fordult. A klasszikus niche-elmélettel összefüggő gyakorlati kérdések arról, hogy mi hová ültethető és hol terem jól, újabban kiegészültek az adaptációs formák sokféleségének és téridőbeli szerveződésének funkcionális következményeire vonatkozó kérdésekkel.
Az állomány léptékű diverzitás a korábbiaknál jobban interpretálható, ha figyelembe vesszük, hogy a fajok az adott élőhelyen belül a forrás- és kondícionáló tényezőknek nem az átlagához, hanem ezek téridőmintázataihoz adaptálódnak. Mérések igazolják, hogy a forrás- és a kondicionáló tényezők állományon belül is jelentősen variálnak, értékük a növényegyedek mérettartományában is folyamatosan változik. Az új paradigma szerint a populációk ezekhez a dinamikus környezeti mintázatokhoz adaptálódnak, illetve visszacsatolások révén azokat befolyásolják. Új eredmény, hogy a források mintázataihoz a populációk növekedési és terjedési stratégiákkal is adaptálódnak (Oborny, 2001). A hasonló viselkedésű, a forrás és a kondicionáló tényezőkhöz hasonló módon adaptálódó és a társulásban hasonló szerepet betöltő fajokat ún. funkciós csoportokba soroljuk (például egyévesek, évelők, sótűrő vagy szárazságtűrő fajok). Az új, nem-egyensúlyi szemlélet elterjedésével az elmúlt években jelentősen kibővült a funkcionális attribútumok köre. A környezeti tényezők mintázataira adott válaszokkal kapcsolatosan fontossá (azaz funkcionálisan interpretálhatóvá) váltak a populációk térbeli elhelyezkedésével, az egyedek méretével, alakjával, integrációjával, illetve ezen tulajdonságok időbeli változásaival (dinamikus morfológia) kapcsolatos attribútumok is.
A funkcionális vegetációtan fejlődésében, módszereinek egységesülésében jelentős szerepet játszanak az ökoszisztéma -modellek új változatai, az ún. egyedalapú mechanisztikus szimulációs modellek. Segítségükkel részletesen megjeleníthető a növényegyedek viselkedése (például csírázás, növekedés, interakciók, szaporodás, halál) és sokezer egyed együttes követésével a korábbiaknál lényegesen részletesebben és realisztikusabban reprezentálható az állományok dinamikája. A vegetációt képviselő (szub)modell más, az ökoszisztéma egyéb komponenseit (például a talajnedvesség téridődinamikáját) megjelenítő modellekkel köthető össze. E nagyon bonyolult modellek eredetileg konkrét (gyakorlati szempontból kiemelten fontos) állományok részletes leírására születtek. Az új, egyszerűsített változatok, amelyeket a globális változások modellezése érdekében fejlesztettek, már különböző vegetációtípusok összehasonlítására is alkalmasak. Az az igény, hogy e modellek input paramétereit távoli és különböző összetételű társulásokban összehasonlítható módon becsülhessük, nagyban hozzájárult a szabványosított mérési eljárások kialakulásához. A legújabb kutatási projektek olyan multinacionális, interkontinentális vállalkozások, amelyek valamilyen általános kérdés (például a szervesanyag-dekompozíció hőmérsékletfüggése, a különböző dinamikai állapotú közösségek invázióval szembeni érzékenysége vagy a rövid életű fajok szerepe a szemiarid rendszerekben) vizsgálata során nagyon sok és nagyon különböző vízi és terresztris rendszer párhuzamos mérésével már általánosítható, globálisan is robusztus és releváns összefüggések felismerését teszik lehetővé.
A funkcionális vegetációtan legfontosabb kérdései ma a globális ökológiai változásokkal kapcsolatosak. Kérdés, hogy a globálizálódó ipar és kereskedelem hatására, a földrajzi izoláció jelentős csökkenésével milyen további fajok terjednek el, ezek képesek-e beépülni a helyi növényközösségbe, vagy túlszaporodva, olyan mértékig alakítják át a lokális környezet téridőmintázatát, hogy kiirtják a korábbi közösséget. Kérdés az is, hogy milyen funkcionális következményekkel jár egy-egy faj elvesztése. Arra is keressük a választ, hogy megváltozott körülmények között milyen növényzeti típusok képesek önnfenntartó módon tovább működni, illetve szerkezeti változások esetén hogyan változik a stabilitás, a növény-állat-mikroorganizmus kapcsolatok és a vegetáció szerepe az anyag- és energiaforgalomban.
A diverzitáscsökkenés funkcionális következményeinek a kutatása az elmúlt tíz év legforrongóbb és leggyorsabban fejlődő területe (Loreau et al., 2001). A 70-es években indultak azok a terepi körülmények között végzett hosszú távú vizsgálatok, amelyek párhuzamosan rögzítették a fajkompozíció, a diverzitás, a növényi produkció és a környezeti paraméterek változásait. A hosszú távú adatsorok feldolgozása során bizonyították először, hogy a diverzebb közösségek produkcióját kevésbé zavarja meg egy-egy környezeti perturbáció (például aszály), illetve a zavarást követően funkcióik (például szervesanyagprodukciójuk) gyorsabban helyreállnak. A mérések azt is kimutatták, hogy azonos körülmények között a kisebb diverzitású rendszerek kevésbé efficiensen hasznosították a forrásokat. Tíz évvel az első eredmények után a kép artikuláltabb, és a diverzitásfunkció-összefüggés különböző típusairól beszélhetünk. Eközben az ökológiai rendszerek diverzitásának folyamatos csökkenése mint globális jelenség, egyre jobban dokumentált, vitathatatlan tény. Sokasodnak azok a vizsgálatok is, amelyek az ökológiai rendszerek funkcionális zavarairól, valamely ökológiai szolgáltatás sérüléseiről számolnak be.
Mely természettudósok
a Természet igazi tudósai?
Mindez közelebb visz a legvégső kérdéshez: kijavíthatók-e a leromlott, diszfunkciós ökológiai rendszerek, lecserélhetők-e a természetes ökológiai rendszerek hasonló vagy jobb hatásfokkal működő mesterséges rendszerekre? A funkcionális vegetációtan eredményei alapján megtervezhető és létrehozható-e egy terület optimális növényzete? Megoldhatók-e a környezeti problémák a biotechnológia segítségével?
A reneszánsz óta a modern tudomány sikert sikerre halmozva hoz létre fantasztikusnál fantasztikusabb technikai újításokat. E folyamat lényege, hogy az emberi populáció természetes környezetét egyre nagyobb mértékben mesterséges környezetté alakítja. A módszer egyelőre példátlanul sikeresnek tűnik, aminek objektív bizonyítéka a humán populáció méretének példátlan növekedése és az ökológiai források fölötti példátlan dominanciája. Mára a szárazföldre jutó bioszféra felét alkotják többé-kevésbé mesterséges, emberkéz alkotta ökológiai rendszerek. Kérdés, hogy van-e, és hol van ennek a folyamatnak a határa? Rendelkezik-e az emberiség azzal a szemlélettel és kultúrával, ami képessé teheti saját környezetének mesterséges kialakítására, fenntartására és újratermelésére? Redukálható-e a földi bioszféra egyetlen faj környezetére? Sikerülhet-e a technika segítségével túlszárnyalni a természetes ökológiai rendszerek teljesítményét? Jelenlegi tudásunk alapján ez eddig még soha nem sikerült. Az ökológiai evidenciák szerint az emberi beavatkozások az ökológiai rendszer bizonyos (rendszerint egyetlen) komponensét szaporítják fel, miközben a rendszer egészének a teljesítménye romlik. Tudjuk például, hogy a szántóföldi monokultúrák és a homogén faültetvények fenntarthatósága véges, a kultivált populációk genetikailag leromlanak, a talaj tápanyag-újratermelő és -megtartó képessége csökken, a mesterséges ökológiai rendszerekben gyakoribbak a predátorok és patogének gradációi, és a monokultúrák kevésbé állnak ellen az időjárás viszontagságainak (Somogyi, 2001; Vida, 2001).
A technológiai megoldás termékekben gondolkodik, amelyet gyárak állítanak elő. A jelenleg uralkodó felfogás szerint a tökéletes termék ismérve a homogén minőség, a tökéletes gyáré pedig a megtermelt mennyiség állandó növelése. Az ökológia törvényei szerint egy hasonló jelenség a szupraindividuális rendszerek körében maga az ökológiai katasztrófa. A nem-egyensúlyi paradigma hívta fel a figyelmet arra, hogy természetben is előfordulnak lokális szabályozási katasztrófák. Ez rendszerint valamilyen zavarás hatására következik be, amikor is egy populáció a többiek rovására elszaporodik, és monopóliumot szerez a források felett. Érdekes módon ez az állapot nem marad fenn stabilan, hanem idővel a monopóliumot szerzett populáció is összeomlik, és helyén valamilyen diverz közösség regenerálódik. A hasonlatnál maradva, a tapasztalat azt mutatja, hogy az evolúció során azok a természetes "ökológiai gyárak" bizonyultak életképesnek, és azoknak sikerült legyőzni versenytársaikat, amelyek egyszerre sok egymással összefüggő terméket gyártottak, a gyártás során állandóan adaptálódtak a piac követelményeihez, és ahol ugyanazon gyáron belül megvalósult a termelőeszközök karbantartása és újratermelése, valamint az innováció és a hulladékgazdálkodás is. Ha az emberiség a jövőben arra kényszerül, hogy maga állítsa elő saját környezetének elemeit, a számára nélkülözhetetlen forrás- és kondicionáló tényezőket (például a belélegezhető levegőt, az iható vizet, az elviselhető klímát és az élelmiszereket), vajon alkalmazhat-e olyan technológiát, ami ellentétes az evolúció és az ökológia törvényeivel? Az ökológia törvényei felől nézve a humán populáció jelenlegi "sikertörténete" már valószínűleg az ökológiai katasztrófa első felvonása.
A funkcionális szemléletű vizsgálatok legfontosabb konklúziója, hogy a szupraindividuális rendszerek diverzitása és rendezettsége a normális működés szükséges feltétele. Ebből következik, hogy azok az ún. mikrokozmosz-kísérletek, amelyek kis területen, rövid ideig, homogén környezetben, csupán néhány faj néhány egyedének bevonásával folynak, a megérteni kívánt mintázati jelenséget nem reprezentálhatják megfelelően. Ezért eredményeiket csak nagyon óvatosan szabad interpretálni, és arra semmiképpen sem alkalmasak, hogy segítségükkel az ökológia nagy (a földi evolúciót befolyásoló) kérdéseit (mint például a természetes és a mesterséges ökológiai rendszerek fenntarthatóságának feltételei) megválaszoljuk. Az új ökológiai paradigma azzal, hogy felhívta a figyelmet a mintázati jelenségek fontosságára és az ebből következő léptékfüggésre (beleértve a nagy tér- és időléptékű jelenségeket is) alapvetően átalakította az ökológiai kísérletek módszertanát.
Kutatások
biológiailag relevánsabb léptékekben
A biológiailag releváns léptékekhez való alkalmazkodás igénye hívta életre a hosszú távú ökológiai kutatásokat (Kovács-Láng - Fekete, 1995). Az Egyesült Államokban az 1970-es évek közepére kiderült, hogy az IBP és a MAB programok által feltett kérdések a programok eredetileg tervezett időkeretében nem válaszolhatók meg. Világossá vált, hogy az addigi mérések térben túl lokálisak, időben pedig túlzottan rövidek és esetlegesek, emiatt az eredmények nem terjeszthetők ki a gyakorlat és a társadalom számára releváns regionális léptékre. A probléma megoldására olyan kutatásfinanszírozási rendszert hoztak létre, amely túlmutatott a korábbi egy-három éves pályázati időszakokon, valamint létrehozták azt az infrastruktúrát, amely egy-egy biomon belül egy-egy tipikus (sok ezer hektáros) nagy területen biztosította a hosszú távú, tervezetten több évtizedes munka biztonságos feltételeit. A területeken belül folyamatosan és automatikusan regisztrálják a környezeti paramétereket, és szervezett módon, táji léptékben és időben hosszú távon kezelési kísérleteket folytatnak. A kezeléseket az adott biomra jellemző természetes zavarási rezsim figyelembevételével alakították ki. A hosszú füvű préri (Konza Prairie LTER hosszú távú ökológiai kísérleti terület, vö. Knapp et al., 1998) esetében például a kezelések a különböző gyakoriságú égetéseket jelentik, amit marhákkal vagy bölényekkel való legeltetéssel kombinálnak. A kezelések egysége egy-egy völgyrendszer (több száz ha), azaz a táj egy természetes funkcionális egysége, elegendően nagy terület ahhoz, hogy az eredmény a gyakorlat számára is közvetlenül hasznosítható legyen. A kezelések folyamatos végrehajtását és a terület őrzését a technikai személyzet végzi. A területen a kutatók számára szálláslehetőség és egyes gyorsan elvégzendő mérésekhez, illetve számításokhoz laboratórium áll rendelkezésre. Az így kiépített területen sokféle szakirányú kutató dolgozik és végzi saját speciális vizsgálatait, amelyeket azonban összekapcsol a közös terület és a közösen vizsgált kezelési kísérlet. A konkrét kutatások különböző léptékben folynak (a távérzékeléssel reprezentált táji léptéktől az egyetlen levélen végzett ökofiziológiai mérésekig), és abban különböző (szub)diszciplínák kutatói (ökoszisztéma-, társulás- és populációökológusok, talajtanosok stb.), illetve különböző élőlénycsoportok szakértői (botanikusok, zoológusok, mikrobiológusok) vesznek részt. Az egyes kutatási eredmények összekapcsolását segítik a területre kidolgozott szimulációs modellek, amelyek optimális esetben hierarchikusan szervezettek (a térinformatikai modellektől az egyed alapú vagy az ökofiziológiai modellekig).
Az itt vázolt kutatási módszertan az Egyesült Államokban szervezetten húsz éve folyik, de az elmúlt tíz évben az egész világon elterjedt, és lassan általánosan elfogadott szabvánnyá válik. Fontos leszögezni, hogy a vegetációdinamikai kutatások jelenlegi felépítését az ökológiai rendszerek tulajdonságainak fokozatos megismerése tette szükségessé, azaz a komplexitásukról, hierarchikus szerveződésükről, a környezeti és a toleranciamintázatok jellemző léptékeiről, és az állatok, valamint az ember szerepéről megismert tényekhez való kényszerű alkalmazkodás.
Hogyan befolyásolhatja a vizsgálat léptéke (például időtartama) a vizsgálat eredményét? Nézzünk néhány példát! Az első esetben arra keresünk választ, hogy mi a szerepe a növénytársulásokban a közepesen gyakori és a ritka fajoknak. A vizsgálat során az egyes kezelésekből rendre kiirtjuk a domináns, a közepesen gyakori, illetve a ritka fajokat, és figyeljük a következményeket. Trópusi gyepek esetében figyelték meg, hogy a ritka fajok eltávolítása után az ott maradó fajok teljesen, a közepesen ritka fajok eltávolítása után pedig részlegesen kompenzálták az állományszintű szervesanyag-produkciót. Viszont ugyanebből a rendszerből a domináns fajt eltávolítva, a produkció lényegesen csökkent. Ha ezt a vizsgálatot nem hónapokig, hanem évekig folytatjuk, már azt tapasztaljuk, hogy a gyep a domináns faj hiányát is képes kompenzálni. A domináns faj kiirtása után a közepesen gyakori fajok részvételével egy, a sebgyógyuláshoz hasonló regenerációs folyamat indul be, és a fajok egymást jellegzetes sorrendben helyettesítve három-öt év után már közel teljesen helyreállítják a gyep funkcióit, akkor is, ha az eredeti domináns faj visszatérésére még éveket kell várni. A közepesen gyakori fajok más alkalmakkor (például az időjárás fluktuációi, herbivor gradációk vagy tűz esetén) is átveszik időlegesen a domináns faj szerepét. Egy újabb elmélet szerint az egészen ritka fajok a több száz vagy több ezer éves időléptékű folyamatokban kapnak szerepet, például a jégkorszakok és az interglaciálisok váltakozása idején. Ez azt is jelenti, hogy az egészen ritka növényfajok szerepének tisztázásához a rövid távú megfigyelések nem adnak érdemi információt, vagy félrevezető eredményre vezethetnek.
A másik példa kapcsán azt vizsgáljuk, hogyan befolyásolja a gyakori égetés a talaj tápanyagkészletét és a legelő fenntarthatóságát a hosszú füvű préri esetében. A préri gyakori (évente való) tavaszi égetése egy-kettő-négy év távlatában (ameddig a hagyományos kutatási projektek általában tartanak) egyértelműen pozitív beavatkozásnak mutatkozott. Az égetés hatására visszaszorultak a gyomok (itt például a Bromus tectorum), megerősödött a domináns őshonos fűfaj, és nőtt a produkció. Ahhoz viszont legalább tíz-tizenöt évig kellett folytatni a kísérleteket, hogy a káros hatások első jelei megmutatkozzanak. Tíz év után a talaj C- és N-tartalma csökkenni kezdett, a gyep érzékennyé vált a szárazságra, felnyílt, beindult az erózió, és csökkenni kezdett a produkció is. Tehát egy kezelés, ami egy-öt év távlatában kedvezőnek bizonyul, még nem biztos, hogy hosszabb időtávon is alkalmazható, fenntartható. Mivel a fenntartható tájhasználat lényegénél fogva hosszú távú feladat, a kezelési tervek kialakításakor nagyon óvatosan kell eljárni. Ott, ahol rövid távú tapasztalatok birtokában tervezik meg egy terület hosszú távú kezelését, a kezelés hatásának léptékfüggése (a kezdetben pozitív hatás későbbi esetleges elmaradása vagy negatívba fordulása) igen komoly probléma, veszélyforrás lehet.
A magyar kutatók a környező országok közül elsőként ismerték fel a modern nem-egyensúlyi paradigma jelentőségét és módszertani konzekvenciáit, és már több mint tíz évvel ezelőtt csatlakoztak a hosszú távú ökológiai kutatások nemzetközi szervezetéhez (ILTER). Az új szemlélet hazai elterjedését elősegítette az a szerencsés tény, hogy Magyarország mind az ökológiai mintázatok leírásában, mind a téridődinamikus modellezésben a világelsők közé tartozik (vö. Juhász-Nagy, 1980; Czárán, 1998). Hazánkban eddig három helyen sikerült megszervezni a hosszú távú ökológiai kutatások nemzetközi szabványának megfelelően a kutatásokat (Balaton Projekt, Síkfőkút Projekt, Kiskunság LTER Projekt). Ezek közül a Kiskunságban a természetvédelemmel szorosan együttműködve, a tájképi mozaik hierarchikus jellegét és sokféleségét is reprezentálva, több kutatóhely közreműködésével és számos diszciplína együttműködésével folynak komplex kutatások (Kovács-Láng - Fekete, 1995; Kovács-Láng et al., 1999). Jelenleg egy Széchenyi-pályázat keretében a természetvédelmi kezelési tervek ökológiai megalapozása a kutatás egyik célkitűzése. A projekt keretében ugyan lokálisan (Fülöpháza és Ágasegyháza térségében), de a táji léptéket reprezentáló nagy területen (több 100 ha) és sokféle aspektusból (populáció- és társulásdinamika, tájökológia, restaurációs ökológia, produkciós ökológia) folynak intenzív, terepkísérletekre alapozott kutatások. Az itt alkalmazott modern módszerek lehetővé teszik, hogy a helyben végzett kutatások más hasonló európai és amerikai kutatóhelyek kutatásaival összekapcsolódva (ILTER Network) a globális (éghajlati és tájhasználati) változásokból adódó kérdésekre is válaszoljanak.
Kitekintés
Hosszú távon, evolúciós perspektívában gondolkodva, egy tudományág annyit ér, amennyire az emberi populáció adaptációját segíteni képes. Az emberiség előbb a csillagokat, majd a mikrovilágot ismerte meg. Sokáig úgy véltük, hogy az előttünk álló nagy problémák megoldásához elegendő a makro- és a mikrovilág törvényszerűségeinek az ismerete. Korunk (újra)felfedezése, hogy a makro- és a mikrovilág között létezik egy, a modern ember számára már rejtett világ, a bioszféra egyedszint feletti szerveződésének és működésének a világa. Létezését sokáig elhanyagoltuk, és jelenlétét ma is inkább csak közvetve érzékeljük, a bioszféra funkcionális zavarain, a környezeti szélsőségek és diszfunkciók (például aszályok, árvizek, szélviharok és járványok) egyre gyakoribb és feltűnőbb jelentkezésén keresztül. Közös érdekünk, hogy még időben megismerjük e rejtett szerkezetek fennmaradásának és normális működésének feltételeit.
A szupraindividuális szerkezetek és folyamatok új, nem-egyensúlyi szemlélete egy reálisabb, ezért a gyakorlatban jobban alkalmazható tudás megszerzésére ad elméleti és módszertani lehetőségeket. A korábbi túlegyszerűsített irányzatokkal szemben egy komplexebb, érdekesebb világ képét ígéri. De a realitásokhoz, azaz a szupraindividuális rendszerek komplexitásához és léptékeihez való alkalmazkodás azzal a következménnyel is jár, hogy a dinamikai folyamatokba való beavatkozáshoz szükséges tudás megszerzése nagyobb szervezést, több munkát és komolyabb infrastrukturális beruházásokat kíván annál, mint azt korábban gondoltuk.
Az új ökológiai paradigma az ökológiai alapkutatások számára igen magasra tette a mércét. Az elvárás az, hogy nagy területen, több évtizedes távlatban tervezhetően, többféle tudományág munkáját operatívan integrálva történjen magas szintű kísérletes munka. A kísérletek arra keresik a választ, hogy melyek azok a legfontosabb mechanizmusok, amelyek az adott ökológiai rendszer hosszú távú fennmaradását és normális működését (az ökológiai szolgáltatásait, a bioszféra egészével fenntartott anyag- és energiaforgalmát) biztosítják. A kísérletek kialakítása során az adott vegetációtípusokat jellemző természetes zavarásokat és a hagyományos emberi tájhasználat hatását igyekeznek utánozni, illetve kicsit átszabni, a megváltozott környezeti feltételeknek megfelelően.
Az ökológusok egy allometrikus összefüggést találtak az állatok testmérete és populációmérete között. Ennek alapján a humán populáció természetes (azaz az eredeti ökológiai kényszereket követő) lélekszáma csak néhány tízmillió lehetne. A vadászó, gyűjtögető ősember populációmérete becslések szerint megfelelt ennek a nagyságrendnek. A kulturális-technikai fejlődés a természet adta korlátokhoz képest hihetetlenül megnövelte az emberiség lélekszámát, egyben kitágította lehetőségeinket a bioszféra szerkezetébe és működésébe való beavatkozásra. Ezeket a változásokat azonban nem követte a környezetünk érzékelésének a megváltozása. A humán populáció ökológiai niche-e megnőtt, azaz a forrás és kondicionáló tényezők topológiai terének egyre nagyobb tartományát használjuk. E növekedés során azonban valahol elvesztettük azt a képességet, hogy érzékeljük az általunk elfoglalt, használt és befolyásolt környezetet. Ma már nem abban a léptékben él az emberiség, amelyben eredetileg kialakult. Veszélyes helyzet állt elő azzal, hogy jelenleg a technikai evolúció gyorsabb, mint kognitív apparátusaink evolúciója. Ennek következménye, hogy elvesztettük a saját territóriumunkban való tájékozódás képességét. Pedig ez a képesség az adaptáció, azaz a fennmaradás alapfeltétele. Korunk tudományának nagy kihívása, hogy helyreállítsa ezt az elveszett képességet. A vegetációtan az általa vizsgált léptéktartományok kitágításával (a növényzeti állományokon belüli mikromintázatok felé, illetve a táji és a globális mintázatok felé), a sokféleség funkcionális szerepének a felismerésével, a mintázatok, mechanizmusok és funkciók operatív összekapcsolásával képessé vált arra, hogy választ adjon a gyakorlati kihívásokra. A vegetációtan specialistái, más diszciplínák kutatóival együttműködve, jelentősen hozzájárulhatnak, hogy visszaszerezzük tájékozódási és adaptációs képességünket mai, gyorsan változó környezetünkben.
Kulcsszavak: degradáció, diverzitás-mintázatok, funkciós csoportok, koegzisztencia feltételek, léptékfüggés, LTER, nem-egyensúlyi vegetációdinamika, ökológiai szolgáltatások, primer produkció
IRODALOM
Bartha Sándor (2000). In vivo társuláselmélet. In: Virágh K. - Kun A. (szerk.) Vegetáció és dinamizmus. MTA-ÖBKI, Vácrátót, 101-141
Bartha Sándor - Czárán Tamás - Podani János (1998). Exploring Plant Community Dynamics in Abstract Coenostate Spaces. Abstracta Botanica. 22, 49-66
Czárán Tamás (1998). Spatiotemporal Models of Population and Community Dynamics. Chapman and Hall, New York
Harper, John L. (1977). Population Biology of Plants. Academic Press, London
Juhász-Nagy Pál (1980). A cönológia koegzisztenciális szerkezeteinek modellezése. Akadémiai doktori értekezés, Budapest
Juhász-Nagy Pál - Podani János (1983). Information Theory Methods for the Study of Spatial Processes and Succession. Vegetatio. 51, 129-140
Knapp, Alan K. - Briggs, J. M. - Hartnett, D. C. - Collins, S. L (eds.) (1998). Grassland Dynamics. Long-term Ecological Research in Tallgrass Prairie. Oxford University Press, N. Y.
Kovács-Láng Edit - Fekete Gábor (1995). Miért kellenek hosszú távú ökológiai kutatások? Magyar Tudomány. 102, 377-392
Kovács-Láng Edit - Molnár E. - Kröel-Dulay Gy. - Barabás S. (szerk.) (1999). Long Term Ecological Research in the Kiskunság, Hungary. KISKUN LTER, MTA-ÖBKI. Vácrátót
Loreau, Michel et al. (2001). Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science. 290, 804-808
Oborny Beáta (2001). A növények térfoglaló és táplálékkereső stratégiái. In: Bartha Zoltán - Liker András - Székely Tamás (szerk.) Viselkedésökológia: modern irányzatok. Osiris, Budapest, 43-62
Pickett, Stuart T. A. - Parker, Victor T. - Fiedler, Peggy Lee (1992). The New Paradigm in Ecology: Implications for Conservation Biology above the Species Level. In: Fiedler, Peggy Lee - Jain, Subodh (eds.) Conservation Biology: The Theory and Practice of Nature Conservation, Preservation, and Management. Chapman & Hall, London, 65-88
Somogyi Zoltán (2001). Erdő nélkül? L'Harmattan, Budapest
Tuba Z. - Csintalan Zs. - Nagy Z. - Szente K. - Kemény G. - Takács Z. - Koch J. - Badacsonyi A. - Murakeözy, P. - Palicz G. - Kóbor Sz. - Ötvös E. - Bartha S. (1998). Szünfiziológia: alapozó gondolatok és exploratív vizsgálatok egy születő növényökológiai tudományterülethez. In: Fekete G. (szerk.): A közösségi ökológia frontvonalai. Scientia, Bp., 171-196
Vida Gábor (2001). Helyünk a bioszférában. Typotex, Budapest
régi: egyensúlyi új: nem-egyensúlyi
társuláselmélet társuláselmélet
Prior textúra-állapotok, zárt téridő mintázatok, téridő
referencia: szimplex konzervatív folyamatok, nyílt, komplex
rendszerek disszipatív rendszerek
Lépték: mesterséges természetes, karakterisztikus
felbontás és kiterjedés skálapontok és intervallumok
Modellek: klasszikus téridő modellek, főként objektum
nem térbeli modellek kölcsönhatás modellek
Törvények: egyszerű univerzális elvek, kontextusfüggő dinamikus
a klasszikus modellek korlátozási relációk
paramétereivel megfogal- a mintázat transz-
mazott egyenlőtlenségek formációk rendje
Mintázat indikál indukál és korlátoz
szerepe:
Értelmező lokális forráshasznosítási ugyanaz, ezenkívül:
populációs efficiencia és a lokális regionális térfoglalási-,
sajátságok: kondicionáló tényezőkkel és terjedési stratégiák,
szembeni tolerancia, térbeli integráció
táplálékkereső stratégiák
1. táblázat * Paradigmaváltás a vegetációtanban. Az egyensúlyi és a nem- egyensúlyi társuláselmélet néhány sajátságának összehasonlítása
1. ábra * A koegzisztenciális mintázattranszformációk alapfolyamatai. A -
regeneráció és degradáció; B - egy hazai növénytársulás (Festucetum vaginatae)
koordináltságának csökkenése a globális ökológiai változásokat reprezentáló
gradiens mentén.
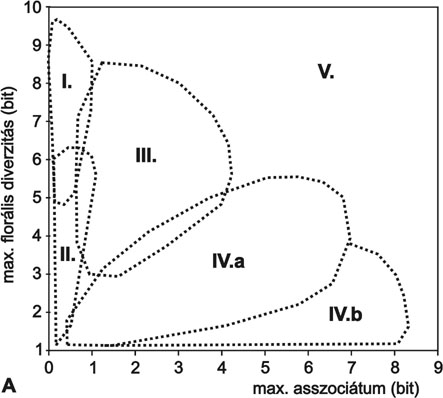
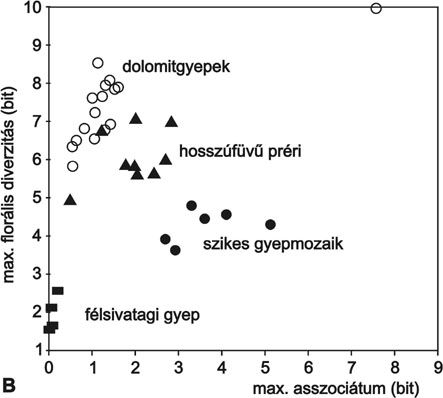 2. ábra * Cönológiai állapotok értelmezése dinamikus neutrális modellekkel. A -
az állapottér felosztása a domináns mintázatképző mechanizmusok szerint (lásd
niche-differenciáció, II. kompetitív dominancia, III. zavarás és kompetitív
dominancia, IV.a és IV.b közepes és erős környezeti heterogenitás, V. tiltott
zóna). B - magyarországi és észak-amerikai növénytársulások elhelyezkedése a
cönológiai állapottérben
2. ábra * Cönológiai állapotok értelmezése dinamikus neutrális modellekkel. A -
az állapottér felosztása a domináns mintázatképző mechanizmusok szerint (lásd
niche-differenciáció, II. kompetitív dominancia, III. zavarás és kompetitív
dominancia, IV.a és IV.b közepes és erős környezeti heterogenitás, V. tiltott
zóna). B - magyarországi és észak-amerikai növénytársulások elhelyezkedése a
cönológiai állapottérben