Ökológia
Scheuring István
a biológiai tudományok kandidátusa, ELTE Növényrendszertani és Ökológiai Tanszék, MTA Ökológiai és Elméleti Biológiai Kutatócsoport
Matematikai modellek az ökológiában
Bevezetés
A XX. század első évtizedei a modern fizika kialakulásának időszaka.
Gyaníthatóan kevesebben tudják, hogy e csodálatos felfedezések árnyékában,
ezidőtájt rakták le a matematikai ökológia alapjait is. Míg a huszadik század
kiemelkedő elméleti fizikusait minden természettudós ismeri, addig az elméleti
ökológia megalapozóiról csak a szakmabeliek hallottak. Emlékezzünk meg tehát
néhány mondatban a nagy elődök munkájáról. Érdekes, hogy többségük (pl. Sir
Ronald Ross, William R. Thompson, Vito Volterra) valamilyen konkrét gyakorlati
probléma megoldásához alkalmaztak matematikai modelleket. Az általános elméleti
kérdésekre koncentráló Alfred J. Lotka inkább kivételnek számított.
A hőskor
Ronald Ross, foglalkozására nézve orvos, sokoldalú tehetséggel megáldott ember
volt. Kifejezett érzéke volt a matematikához, de festett, verseket és
színdarabokat is írt. Orvosként Indiában dolgozott, ahol a malária-fertőzés
ökológiai hátterével foglalkozott. Felismerte, hogy a betegség terjedésének
jellemzői a malária kórokozó, a közvetítő szúnyog és az ember közti kapcsolat
matematikai modelljének segítségével jól magyarázhatók. Dinamikai modelljének
segítségével megmutatta, hogy a betegség csak a terjesztő szúnyogok egy
kritikus denzitása felett marad fenn tartósan, tehát a legjobb védekezés a
szúnyogok irtása. A maláriafertőzés biológiai ökológiai hátterét felfedő
kutatásait 1902-ben Nobel-díjjal jutalmazták.
Vito Volterra a matematikai fizika neves professzora volt Rómában. Figyelmét
ökológiai problémák felé későbbi veje, a biológus Umberto D'Ancona fordította.
D'Ancona az Adriai-tenger halászati adatait elemezve észrevette, hogy az I.
világháború idején, amikor az aknazár miatt a halászat jelentősen visszaesett,
egyes ragadozó halak fogási aránya megnőtt, míg a ragadozók potenciális
zsákmányainak számító halfajok egyedszáma visszaesett. Kimutatta továbbá, hogy
mind a ragadozó, mind a zsákmány halak populációinak mérete egymást követve
ciklikusan változik. Vajon miért van ez így? - kérdezte az ifjú biológus
apósától. Volterra mint igazi fizikus a probléma lehető legegyszerűbb
matematikai modelljét vizsgálta: egyetlen ragadozó és egyetlen zsákmány faj
denzitásának változását egy kétváltozós közönséges
differenciál-egyenletrendszerrel írta le. Modellje segítségével könnyen
magyarázni tudta veje megfigyeléseit. Eredményeit 1926-ban publikálta olasz
nyelven. E cikkről a Nature rövid ismertetőt közölt. Érdekes módon egy másik
fizikusnak, az amerikai Alfred J. Lotkának, egy évvel korábban könyve jelent
meg A fizikai biológia elemei (Elements of Physical Biology) címen, melyben
Volterra eredményei lényegében mind megtalálhatóak voltak. Ez a felismerés az
egyébként elég nehéz körülmények között és szakmai izoláltságban élő Lotkát
meglehetősen felkavarta. Ráadásul könyvének célkitűzése, az ökológiai
kapcsolatok matematikai leírása, sokkal általánosabb volt, mint Volterra
idézett cikkének motivációja. Bár a két tudós viszonya a prioritási viták miatt
később sem volt felhőtlen, a köztük lévő verseny és tehetségük eredményeként
kiemelkedő szerepük volt a matematikai ökológia megteremtésében. Egyikük egy
konkrét problémán keresztül jutott el a legáltalánosabb elméleti kérdések
matematikai megfogalmazásáig, másikuk alapvetően ezzel a szándékkal kezdett az
ökológiával foglalkozni.
Az ökológiai modellek jellemzői
A többi természettudományos diszciplínához hasonlóan a matematika szerepe az
ökológiában a téridő folyamatok tömör leírása, a mintázatok mögött húzódó
mechanizmusok felderítése. Azonban a fizikai vagy a kémiai rendszerekkel
összehasonlítva az ökológiai rendszerek lényegesen összetettebbek. Egy
populáció egyedei különböznek egymástól életkorban, öröklődő és szerzett
tulajdonságokban, továbbá az élettelen környezet legtöbbször térben és időben
is nagymértékben heterogén. Egy élőközösségben számos, sokszor több száz
különböző populáció egyedei vannak egymással kölcsönhatásban, s ezek a
kapcsolatok jellemzően nemlineárisak és sztochasztikusak. Ráadásul, ami fontos
lehet például egy vízi élőközösségben, az egyáltalán nem lényeges mondjuk egy
gyeptársulásban vagy egy erdőben. Míg például egy gyeptársulás kialakításában
döntő szerepe van a térbeli kényszereknek, addig a planktonközösségekre inkább
az áramló víz fizikai tulajdonságai vannak nagy hatással. Maga a vizsgálat
tárgya rendkívül diverz és komplex, felépítése alapvetően hierarchikus.1 Ebből
következik, hogy a rendszereket leíró matematikai modellek is nagyon sokfélék
és bonyolultak. Igen ám, de ezekből a nagyon részletes, sokparaméteres és
sokváltozós matematikai modellekből általános következtetéseket aligha lehet
levonni. Ezt a problémát felismerve, az elméleti ökológusok kutatásaik során
két irányt követnek. Az első irányzat (hasonlóan Lotkához és Volterrához)
igyekszik minél egyszerűbb, a folyamat lényegét megragadó ún. stratégiai
modellek segítségével minél szélesebb körben érvényes megállapításokat tenni, a
második csoport egy-egy működésében jobban ismert rendszert igyekszik minél
pontosabban leírni ún. részletes modellekkel (ahogy egykor Ross is tette). Az
első csoport inkább alap-, a második inkább alkalmazott kutatásokkal
foglalkozik.
Természetesen a fizikai vagy kémiai modellekhez hasonlóan az ökológiai modellek
is alapvető posztulátumok alapján építik fel a konkrét modelleket. A
legfontosabb azt leszögezni, hogy az ökológiai kutatások alapegysége a
populáció, bár sokszor érdemes (és érdekes) a populációnál magasabb
szerveződési szintű egységek (élőközösségek, társulások, biomok) dinamikai
viselkedését vizsgálni. Mivel a populációkról és az azokat felépítő egyedekről
többet tudunk, mint a magasabb szerveződési egységekről, általános érvényű
posztulátumokat a populációkra lehet megfogalmazni. Az egyik legfontosabb
kiinduló posztulátum, hogy ha a környezeti hatásokban nincsenek szisztematikus
változások, akkor a populáció egyedszáma exponenciálisan változik. Ez
egyszerűen azért van, mert ebben az esetben a populáció egyedszámváltozását (az
időátlagban) állandó születési és halálozási ráták határozzák meg. Ez a
megállapítás analóg a newtoni tehetetlenség törvényével: egy teljesen ideális
esetben, melyet igazából csak gondolatkísérlet útján tudunk előállítani, meg
tudjuk mondani a test, illetve a populáció dinamikai viselkedését.
Természetesen az exponenciális növekedés, illetve csökkenés elvét a
megfigyelések és a kísérletek nagyon jó közelítéssel igazolják. Egy táptalajban
gazdag baktériumtelepen a populációméret kezdetben exponenciálisan növekszik,
ilyen a betegségszám növekedése minden járvány kitörésekor is.
Ezek után azokat a posztulátumokat érdemes lefektetni, melyek segítségével a
valós populációk dinamikai viselkedését leírhatjuk. Ilyen posztulátum például,
hogy minden populációban van egy felső határa a denzitásnak, amelyet a
populáció nem léphet túl. Mivel a rendelkezésre álló források végesek, ez a
józan ésszel teljes összhangban lévő megállapítás. Kimondva-kimondatlanul ezt a
feltevést használjuk, amikor a populációnövekedés denzitásfüggését matematikai
modellekben alkalmazzuk. Például a szaporodáshoz tápanyagfelvételre van
szükség, és a szaporodási sebesség ennek a tápanyagfelvételnek monoton növő
függvénye. A tápanyagfelvétellel kapcsolatban két további általános
összefüggést vagy posztulátumot mondhatunk ki: 1.) alacsony tápanyag-denzitás
esetén a tápanyagfelvétel arányos a forrás denzitásával, 2.) magas
tápanyag-denzitás esetén a fogyasztó fiziológiai képességei felső határt
szabnak a tápanyagbevitelre, s így a szaporodási sebességre is. E két utóbbi
posztulátumon alapulnak a speciális esetekre megfogalmazott úgynevezett
funkcionális válaszfüggvények, azaz azok a függvények, melyek a
tápanyagmennyiség és az egységnyi idő alatt felvett tápanyag között teremtenek
kapcsolatot (Turchin, 2003). Bármilyen jellegű matematikai modelljét vizsgáljuk
a populációknak, illetve a populációk alkotta közösségeknek, társulásoknak,
ezeknek a posztulátumoknak eleget kell tenni.
Egy tudomány akkor fejlődik egészségesen, ha a kísérleti (terepi,
laboratóriumi) és az elméleti (matematikai modelleken alapuló) kutatások
ismerik és alkalmazzák egymás eredményeit. Sajnos az ökológiában ezen a
lényeges ponton komoly nehézségek vannak, elsősorban azért, mert az ökológiai
folyamatok többsége olyan lassú és a rendszerek olyan összetettek, hogy a
kísérletezés és a terepi megfigyelések is komoly akadályokba ütköznek.
Világszínvonalú terepökológiához hosszú időre, átgondolt kutatási stratégiákra
és sok pénzre van szükség. Annak ellenére, hogy ma már sok helyen végeznek
gondosan tervezett, hosszú távú ökológiai kísérleteket az elméleti ökológusok
sokszor nem tehetnek mást, mint igyekeznek a meglévő adatokat összegyűjteni,
elemezni, és ezek alapján a modellek jóslatait a terepi eredményekkel
összevetni, továbbá a stratégiai modellek viselkedéséből viszonylag általános
érvényű hipotéziseket megfogalmazni. A következő fejezetben néhány, a
matematikai modellek támogatásával kialakított hipotézisről, illetve egy-egy
olyan kutatási területről kívánok szólni, ahol a kísérleti és terepadatok
matematikai modellek együttes alkalmazása vezetett új eredményekre.
Modellek, hipotézisek, ellentmondások
A modellezés egyik legfontosabb erénye, hogy a modellek alapján egyértelműen
megfogalmazott feltételek mellett határozott, sokszor nagyon éles
következtetéseket lehet megfogalmazni. Ilyen például a kompetitív kizáródás
elve. A tápanyagforrások és a fogyasztók dinamikáját közönséges
differenciálegyenletekkel leírva arra a megállapításra jutunk, hogy a stabilan
egyensúlyban lévő fajok száma nem haladhatja meg a független források számát. E
matematikai eredmény legizgalmasabb vonása, hogy a terepi megfigyelések
egyáltalán nem támasztják alá. George Evelyn Hutchinson, az elmúlt század egyik
legnagyobb ökológusa, már a negyvenes években felfigyelt arra, hogy az együtt
élő fajok száma a legtöbb élőközösségben messze túlhaladja a limitáló források
számát. Az elmélet és a valóság közötti ellentmondás, mint oly gyakran,
rendkívül termékenyen hatott a tudomány fejlődésére. Azóta számtalan
matematikai modell és terepi megfigyelés eredményeként sokkal összetettebben
látjuk ezt a kérdéskört; kiderült, hogy az adott élőközösségtől függően igen
sokféle, a fajok együttélését fenntartó mechanizmus képzelhető el. Például, ha
a tápanyagforrások vagy (belső dinamikai okok miatt) a populációdenzitások
fluktuálnak, esetleg az élőlények túlélése külső hatások miatt időnként
lecsökken, akkor a tartósan együtt élő fajok száma meghaladhatja a független
források számát. Matematikai modellek segítségével sikerült kimutatni azt is,
hogy a fitoplankton élőközösségekben az áramló víz kaotikus keveredése2 is
hozzájárulhat a versengő fajok együttéléséhez. Annak következtében, hogy a
keveredés hatására a tápanyagforrás és a fogyasztók is egy fraktál mentén
mozognak, a tápanyagfelvétel a kis denzitású fogyasztó számára rendkívül
hatékony lesz. Ennek következtében a kevésbé hatékony kompetitor gyorsabban fog
szaporodni, mint a hatékonyabbak, ha elég kis denzitásban van jelen (például
Károlyi et al., 2000; Scheuring et al., 2003). A fitoplankton esetében a
herbivóroknak (a fitplonktont fogyasztó zooplankton) az élőközösség
szabályozásában általában nincs jelentős szerepe, de számos más közösségben
maga ragadozó vagy valamilyen parazita az, aki a versengő fajokat egyensúlyban
tartja, növelve ezzel a közösség fajgazdagságát. Ezen a ponton átléphetünk egy
másik szövevényes területre: a táplálékhálózatok témakörébe.
Miközben a fajok kapcsolatát elsősorban a forrásokért való versengésként fogtuk
fel, elhallgattuk azt a nyilvánvaló tényt, hogy heterotróf fogyasztók esetén a
források maguk is élőlények. Egy-egy élőközösség akár több száz fajból álló
táplálkozási hálózatot alkot. Számos, távlatilag "életbevágóan fontos" kérdés
tehető fel a táplálékhálózatokkal kapcsolatban: Milyen topológiai és dinamikai
tulajdonságokkal rendelkeznek ezek a hálózatok, vannak-e általános érvényű
jellegzetességeik? Hogyan épülnek föl és mennyire stabilak a külső zavarokkal
szemben? Robert May korszakalkotó könyvében (May, 1974) az ilyen általános
problémák megközelítéséhez használt stratégiai modellek segítségével számos
kiindulópontként használható megállapítást tett. Kimutatta például, hogy a
táplálékhálózat komplexitásának növelésével a modellezett hálózat stabilitása
csökken. Akárcsak a kompetitív kizáródás elve esetén ez a következtetés sincs
igazán összhangban a terepi megfigyelésekkel. Tehát a modellek lényeges ponton
élnek túlzott egyszerűsítésekkel. A valósághoz jobban közelítő matematikai
modellek vizsgálatának eredményeként néhány éve arra hívták fel a figyelmet,
hogy a táplálékhálózatban lévő néhány gyenge kölcsönhatás3 jelentősen
fokozhatja a hálózat stabilitását (McCann - Hastings, 1997; McCann et al.,
1998). Bár e sorok írója saját kutatásai alapján kételkedik az iménti
megállapítások általánosságában, íme egy újabb elméleti eredmény, melynek
jelentős szemléletformáló ereje volt az utóbbi években.
Eddig olyan kutatásokról tettünk említést, ahol a modellezés meglepő, a
megfigyelésekkel ellentmondó eredményei sarkalták mind az elméleti, mind a
terepbiológusokat az adott kérdéskör mélyreható tanulmányozására. Vannak
esetek, amikor a rendelkezésre álló adatok elemzése a modellezési ismeretekkel
társulva hoznak létre érdekes felfedezéseket. Az ökológiai rendszerekre
jellemző nemlineáris kölcsönhatások eredményeként a populációk denzitása
gyakran ciklikusan változik. Matematikai modellek segítségével megmutatható,
hogy ha a populáció növekedését nem az aktuális táplálékmennyiség határozza
meg, hanem a populáció demográfiai szerkezete, akkor a ciklus periódusa
rövidebb lesz, mint olyan populációknál, ahol a növekedést egy forrás
limitálja. Ezt az eredményt és a meglévő terepi adatokat felhasználva
nemrégiben kimutatták, hogy a generalista (sok független forrást kiaknázni
képes) fajok dinamikai szempontból úgy viselkednek, mintha egyetlen, a
táplálékhálózatból kiemelt fajt vizsgálnánk, a specialisták viszont hosszabb
periódussal fluktuálnak. Tehát a generalisták csak gyengén csatolódnak a
táplálékhálózatba, ezzel szemben a specialisták a forrásokhoz erősen csatolódva
alkotnak rendszert (Murdoch et al., 2002). Lehet, hogy ennek alapján a
nagyméretű táplálékhálózatok elemzése is egyszerűbbé válik? Vajon szélesebb
adatbázist figyelembe véve, illetve nem-ciklikus esetekben is lehet-e hasonló
megkülönböztetést tenni a generalisták és a specialisták között? Ezek egyelőre
nyitott kérdések, de biztos vagyok benne, hogy nem kell sokáig várni a
válaszokra.
Modellek és adatok
Ahogy a gyümölcsmuslica a genetikusok egyik kedvelt kísérleti élőlénye, úgy az
elméleti ökológusoknak is vannak kedvelt kísérleti rendszereik. Fontos, hogy a
vizsgálandó közösség kevés fajból álljon, könnyű legyen azokat laboratóriumi
körülmények között tartani, és rövidek legyenek a generációs idők. Ilyen
szempontból ideálisak a planktonközösségek és bizonyos rovarok is. (Például a
fitoplankton-zooplankton közösségekben a plankton denzitások ciklikusságát az
akvárium színének ciklikus változása azonnal jelzi.) Az egyik legnépszerűbb
laboratóriumi rovar a Tribolium castaneum (lisztbogár). Egy amerikai csoport
lassan egy évtizede tart ilyen rovarokat ellenőrzött körülmények között.
Folyamatosan mérik a populáció demográfiai adatait, és nagyon átgondolt
modellépítéssel próbálják a populáció dinamikáját minél pontosabban leírni
(Dennis et al., 2001). Részletes, de mégsem túlzottan bonyolult modelljük
nagyon szépen illeszkedik a mérésekhez. Ráadásul, a modell jóslataival teljes
összhangban, az imágók halálozási rátájának változtatásával igen sokféle
dinamikai viselkedést elő lehetett idézni. Például, nagyon nagy halálozási
ráták esetén a populáció egyedszáma kaotikusan változik.
Az elméleti ökológusokat a hetvenes évek eleje óta intenzíven érdekli, hogy az
ökológiai rendszerekben mennyire jellemző a kaotikus viselkedés. Bár a
populációdinamikai, ökológiai modellek sokaságában mutatták ki a káosz
jelenlétét bizonyos paramétertartományokban, a lisztbogár talán az egyetlen
meggyőző kísérleti bizonyíték arra, hogy (legalábbis mesterséges körülmények
között) a káosz a populációk lehetséges dinamikai állapota. A meglévő számos,
de igen rövid populációs adatsor viszont nem igazán alkalmas e kérdés
megválaszolására. Arról nem is beszélve, hogy sokváltozós, zajos dinamikai
rendszerekben alacsonydimenziós determinisztikus káoszról4 beszélni
értelmetlen. Ezért az adatsorokból újabban inkább azt próbálják meg kideríteni,
hogy a mögötte lévő dinamikai rendszer mennyire érzékeny a zajra, milyen erős a
rendszerben periodicitás stb. (Turchin, 2003). Ezek a kutatások felhívják a
figyelmet arra, hogy még a legegyszerűbb stratégiai modelleknek is figyelembe
kell venniük, hogy az ökológiában jellemzően nemlineáris kapcsolatok vannak,
ahol a külső és belső hatások zajos jellege nem elhanyagolható.
Folytonos leromlás, növekvő fluktuációk, hirtelen katasztrófák
Az átlagos éves középhőmérséklet, a szén-dioxid-koncentráció növekedése, a
talaj pH-jának csökkenése, a természetes élőhelyek feldarabolódása egyaránt az
emberi hatások okozta folyamatok. De a természetes szukcesszió hatására is
lassú folytonos változások mennek végbe a mikroklímában és a biotikus
feltételekben is. Vajon ezek a változások hasonlóan lassú és megjósolható
deformációkat okoznak az ökológiai rendszerekben, vagy eredményezhetnek igen
gyors katasztrofális változásokat is? A kérdés korántsem akadémikus, de a
megalapozott válaszhoz szükség van a modellezők stratégiai modelljeire. Az
imént hangsúlyozott nemlineáris kölcsönhatások miatt könnyen előfordulhat, hogy
egy ökológiai rendszernek több alternatív stabil egyensúlyi állapota van.
Megfigyelhető ez a jelenség például egy olyan ökológiai rendszer matematikai
modelljében, ahol valamilyen rovar fogyasztja egy erdő lombját, és a rovarokat
egy állandó egyedszámú, aktív keresőképpel rendelkező ragadozó fogyasztja. A
lombkorona biomasszája a lassan változó paraméter a modellben. Az 1. a ábra egy
ilyen rendszer stabil és instabil állapotait mutatja be valamilyen lassan
változó külső paraméter függvényében. Látható, hogy egy adott
paramétertartományban a rendszernek két stabil egyensúlyi állapota is lehet. E
két állapot vonzó tartományát egy instabil állapot választja el egymástól
(szaggatott vonal). A paraméter növelésével az ökológiai állapot folytonosan
változik, mígnem elérkezünk a B1 bifurkációs ponthoz, ahol hirtelen ugrik át a
másik stabil állapotba. Érdemes hangsúlyozni, hogy semmi sem jelzi előre azt,
hogy a rendszer közeledik a bifurkációs ponthoz! Viszont, ha vissza szeretnénk
vezetni a rendszert az eredeti állapotba, akkor a környezeti változó értékét
csökkenteni kell, egészen addig, míg a B2 pontba jutunk. A rendszerben
hiszterézis van, azaz az eredeti állapot visszaállításához nem elégséges, ha
környezeti paramétereket az eredeti szintre állítjuk vissza. A bifurkációs pont
felé közeledve a stabil állapot vonzó tartománya egyre szűkül, tehát a
természetes fluktuációk az ökológiai rendszerben egyre nagyobb eséllyel
okozhatnak drámai változásokat. A rendszer a másik stabil állapotba ugrik át,
ha az ökológiai állapot annyira fluktuál, hogy az instabil állapoton átjutva a
másik stabil állapot vonzáskörébe jut (1. b ábra, F1 pont). Nemcsak a folytonos
környezeti változások, hanem a megváltozott viszonyok miatt megnövekedett
fluktuációk is okozhatnak katasztrofális változásokat (1. b ábra, F2 pont).
Természetesen ezek csupán a matematikai modellekből származó megállapítások, de
a megfigyelések alátámasztják őket. A már említett erdő-rovar-rovarevő
rendszerben közismert példák vannak arra, hogy a lombozat növekedésével a
rovarok egyedszáma lassan, majd egy kritikus értéket elérve ugrásszerűen
növekszik (gradáció). A gradációt a tápnövény letarolása miatt
egyedszámcsökkenés követi, és a ciklus kezdődik elölről. Több sekély tóban az
eutrofizáció is gyaníthatóan hasonló jelenség. Ha a bevitt tápanyagmennyiség
elér egy kritikus értéket, a tóban hirtelen szaporodnak el bizonyos algák, vagy
a hínárállomány tömege növekszik ugrásszerűen (ez persze csak az 1-2 méter mély
tavakban fordul elő). Ettől a tó mélyebb részére nem jut el elég napfény. Ezért
az ott élő fotoszintetizáló élőlények többsége kipusztulhat, majd az őket
fogyasztó állatok is eltűnhetnek a tóból. Az eredetihez hasonló fajkészletű
állapot visszaállításához gyakran a kritikus értéknél jóval alacsonyabb
tápanyagbevitelre van szükség (Scheffer et al., 2001). Tehát a hiszterézis
egyértelműen megfigyelhető.
Összefoglalás
A természettudományok a matematika nyelvén íródnak. Természetesen az ökológia
ebből a szempontból sem kivétel, bár a folyamatok matematikai modellezése
egyelőre szerényebb sikereket képes felmutatni, mint az élettelen
természettudományok. Véleményem szerint ennek legalább három oka van: egyrészt
a vizsgálat tárgya rendkívül összetett, másrészt a kísérletezés és terepi
adatgyűjtés is számos nehézségbe ütközik, harmadrészt ez a tudományterület
rengeteg új matematikai módszer kifejlesztését is igényli.
A nehézségek ellenére a fejlett statisztikai eljárások mellet a matematikai
modellalkotás alapvető és egyre nagyobb fontosságú részét képezi a biológiának
és ezen belül az ökológiai kutatásoknak is. Természetesen hazánkban is vannak
kutatók, akik kifejezetten ökológiai modellezéssel foglalkoznak. Ilyen jellegű
kutatások elsősorban, az ELTE Növényrendszertani és Ökológiai Tanszékén, a
Genetika Tanszékén, a Fizikai Biológia Tanszékén, a Vácrátóti Botanikai
Intézetben és a KLTE Ökológia Tanszékén folynak. Bár csupán tucatnyi
szakemberről van szó, túlzás nélkül állítható, hogy a magyar elméleti ökológiai
iskola eredményeivel nevet szerzett a tudományos világban. A sikerek hátterében
egyértelműen ott található néhai Juhász-Nagy Pál (pl. Juhász-Nagy, 1986),
akinek szellemi hagyatéka és személyes varázsa máig érezhetően éltető ereje az
ökológiai modellezőknek.
Köszönetnyilvánítás
A szerzőt az OTKA 032423-as és a 037726-os számú pályázata, valamint a Bolyai
János kutatói ösztöndíj támogatja. Köszönöm Bartha Sándor, Jordán Ferenc és
Pásztor Erzsébet kritikai észrevételeit, javaslatait.
Kulcsszavak: komplexitás, nemlineáris dinamika, populáció, sztochasztikus
folyamatok
1 Az egyedek populációkat, a populációk ismétlődően tartósan együtt előforduló
populáció-kollektívumokat alkotnak.
2 Kaotikus keveredés esetén a vízben mozgó részecskék pályája kaotikus, bár a
sebességtér az idő folytonosan változó függvénye (ellentétben a turbulens
áramlásokkal). A keveredés erős, de nem tökéletes.
3 Két faj akkor van trofikus kölcsönhatásban, ha az egyik fogyasztja a másikat.
Annak erősségét leggyakrabban az egységnyi idő alatt felvett és leadott
biomassza mennyiségével szokták definiálni. A hatás akkor tekinthető gyengének,
ha az adott populáció többi trofikus kölcsönhatásánál sokkal kisebb.
4 A káosz kifejezést olyan teljesen determinisztikus rendszerekben
megfigyelhető szabálytalan mozgásra alkalmazzák, ahol a független változók
száma (azaz a rendszer dimenziója) nem több, mint hat vagy mondjuk tizenkettő.
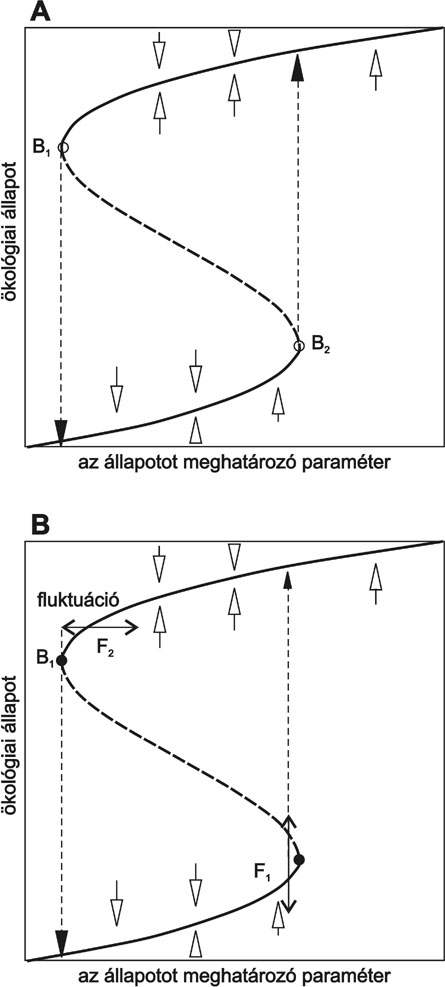 1. ábra * Az egyensúlyi állapotok az ökológiai rendszer valamilyen
paraméterének függvényében. A stabil állapotokat folytonos, az instabilt
szaggatott vonallal jelöltük. a) Ha a paraméter változása során a rendszer
eléri a B1 (B2) bifurkációs pontot, akkor az átugrik a másik stabil állapotba.
b) A bifurkációs pont közelében, az ökológiai állapot fluktuációjának hatására
(F1), a két stabil állapotot elválasztó instabil állapotot átlépve jut el a
rendszer az alternatív stabil állapotba. Az állapotot meghatározó paraméterek
megnövekedett fluktuációja (F2) is vezethet bifurkációhoz.
1. ábra * Az egyensúlyi állapotok az ökológiai rendszer valamilyen
paraméterének függvényében. A stabil állapotokat folytonos, az instabilt
szaggatott vonallal jelöltük. a) Ha a paraméter változása során a rendszer
eléri a B1 (B2) bifurkációs pontot, akkor az átugrik a másik stabil állapotba.
b) A bifurkációs pont közelében, az ökológiai állapot fluktuációjának hatására
(F1), a két stabil állapotot elválasztó instabil állapotot átlépve jut el a
rendszer az alternatív stabil állapotba. Az állapotot meghatározó paraméterek
megnövekedett fluktuációja (F2) is vezethet bifurkációhoz.
IRODALOM
Dennis, Brian - Desharnais, R. - Cushing, J. M. - Henson, S. M. - Costantino,
R. F. (2001). Estimating Chaos and Complex Dynamics in an Insect Population.
Ecological Monographs. 71, 277-303
Juhász-Nagy Pál (1986). Egy operatív ökológia hiánya, szükségszerűsége és
feladatai. Akadémiai, Budapest
Kingsland, Sharon E. (1995). Modelling Nature. The University of Chicago Press,
Chicago
Károlyi György - Péntek Á. - Scheuring I. - Tél T. - Toroczkai Z. (2000). Open
Chaotic Flow: The Physiscs of Species Coexsitence. Proceedings of the National
Academy of Sciences of the USA. 97, 13661-13665
May, Robert M. (1994). Stability And Complexity in Model Ecosystems. Princeton
University Press, Princeton, USA (Second Edition)
McCann, Kevin - Hastings, Alan (1997). Re-Evaluating the Omnivory-Stability
Relationship in Food Webs. Proceedings of the Royal Society of London. Series
B. 264, 1249-1254
McCann, Kevin - Hastings, Alan - Huxel, Gary R. (1998). Weak Trophic
Interactions and the Balance of Nature. Nature. 395, 794-798
Murdoch, William W.- Kendall, B. E. - Nisbet, R. M. - Briggs, C. J. - Mccauley,
E. - Bolser, R. (2002). Single-Species Models for Many Species Food Webs.
Nature. 417, 541-543
Scheffer, Marten - Carpenter, S. - Foley, J. A. - Folke, C. - Walker, B.
(2001). Catastrophic Shifts in Ecosystems. Nature. 413, 591-596
Scheuring István - Károlyi Gy. - Tél T. - Torockai Z. (2003). Competing
Populations in Flows with Chaotic Mixing. Theoretical Population Biology. 6,
77-90
Turchin Peter (2003). Complex Population Dynamics: A Theoretical/Empirical
Synthesis. Princetone University Press, Princeton
<-- Vissza a 2004/1 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]