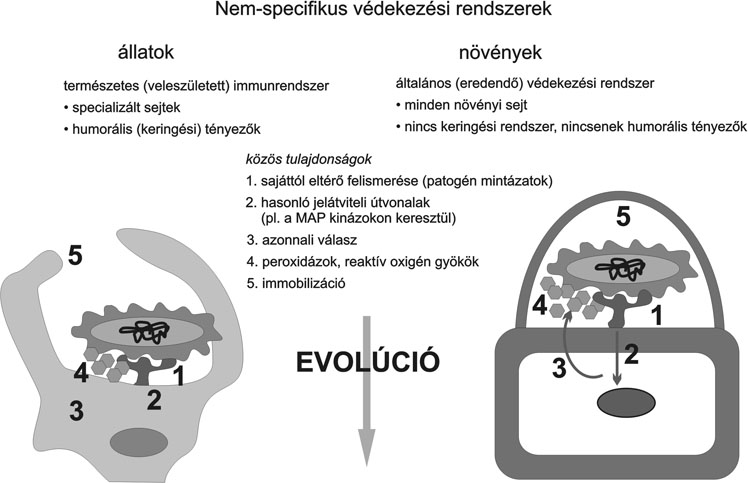
 1. ábra * Védekezés az élővilágban, az állati és növényi védekezés közös és
eltérő tulajdonságai
1. ábra * Védekezés az élővilágban, az állati és növényi védekezés közös és
eltérő tulajdonságai
Mikroorganizmusok népesítik be környezetünket, ha nem is látjuk őket. Csak akkor kezdünk odafigyelni, amikor különböző betegségek, járványok pusztítanak világszerte. Megbetegszünk mi magunk is, elhullanak állataink, és elpusztul az ígéretes termés. A mikrobák nagy része az emberi, állati és növényi szervezetre "ártalmatlan" szaprofiton, másik részük az élőlényekre specializálódott kórokozók. Mindezekkel szemben az élő szervezetek aktívan védekeznek egész életük során. A szaprofitonokkal szemben kialakult természetes vagy általános védekezési rendszer működése során a növényeknél nem jelentkeznek tünetek, így legtöbbször nem is veszünk róluk tudomást. Patogénekkel szemben az általános rezisztencia gyakran már nem elegendő, ezért a védekezésnek egy fejlettebb formája alakult ki, amely már külsőleg is mutatkozó gyenge tünetekkel jelentkezik. Ha ebben a harcban a kórokozó felülkerekedik, akkor a betegség súlyos formái alakulnak ki (Klement, m. a.).
A mikroorganizmusok támadásától a magasabbrendű élőlényeket első vonalban bőrszövetük védi meg. Az ép barriert (hámszövet, növényeknél viaszos epidermisz) csak egyes gomba kórokozók tudják közvetlenül áttörni. A mikroorganizmusok legtöbbször csak sebzéseken vagy természetes nyílásokon át lépnek be a szervezetbe. Növényeknél elsősorban a nyitott légzőnyílásokon vagy a hidatódákon keresztül, de gyakran a rovarok által keltett sebzéseken, szúrásokon át is.
Hajlamosak vagyunk arra, hogy csak a kórokozók indukálta folyamatokkal foglalkozzunk, pedig a védekezési rendszereknek nemcsak a patogéneket kell legyőzniük, hanem a nem kórokozókat (szaprofitonokat) is, hiszen a különböző mikroorganizmusok tengerében élünk, amelyek mindig és mindenhol jelen vannak. Elgondolkoztató, hogy mi történne akkor, ha ezek az ártalmatlannak tűnő szaprofiton mikroorganizmusok táplálékot keresve elárasztanák szöveteinket, és az immunrendszer nem semmisítené meg őket még a megtelepedés helyén. Valóban ez történik akkor, amikor az élet "elszáll", és így a mikroorganizmusok gátlástalanul felszaporodva, órák vagy napok alatt dezorganizálják az elpusztult szervezetet. Hasonló, de nem ilyen gyors folyamat játszódik le, amikor az élő szervezet immunrendszere - legyen az állat vagy növény - valamilyen káros hatás miatt (például lehűlés, hiányos táplálkozás, stresszhelyzetek stb.) legyengül. Erre vonatkozóan jó példákat találunk Bertók Lóránd írásában is. A növényvilágban is ismerünk ún. opportunista patogéneket, amelyek csak ilyen körülményekre várnak, hogy elárasszák a szöveteket, és súlyos betegségeket okozzanak. Ide tartozik például a Pseudomonas syringae pv. syringae baktérium, amely csak lehűlések alkalmával - amikor az általános rezisztencia korlátozódik - okoz betegséget (Besenyei et al., 2003).
A növényi és állati szervezet védekezési rendszerének alapvető azonossága
Mind a növényi, mind az állati immunrendszer lényege abban áll, hogy képes megkülönböztetni egymástól a saját és a nem saját (vírus, baktérium, gomba stb.) anyagokat illetve sejteket, és a felismerés után az idegent hatástalanítani. Minden olyan esetben, amikor ez a felismerés késlekedik vagy elmarad, a betegség súlyos formája jelentkezik. A növények esetében is, az állati immunrendszerhez hasonlóan, kétféle védekezési rendszert különíthetünk el: (1) Általános (eredendő) rezisztencia, ami az állatvilágban természetes (veleszületett) immunitásnak felel meg; (2) a kórokozókra fajlagos ún. specifikus (hiperszenzitív) rezisztencia (HR), amely az állatvilágban az adaptív (szerzett) immunrendszerrel analógnak tekinthető (1. ábra). Még mielőtt ezeket a védekezési mechanizmusokat összehasonlítanánk, fel kell hívni a figyelmet a növényi és állati szervezet funkcionális jellegéből adódó alapvető különbözőségére is (Szatmári - Klement, 2003).
A növényi és állati szervezet védekezési rendszerének alapvető különbözősége
A növényi és az állati védekezési rendszer jelentősen eltér abban, hogy míg minden egyes megtámadott növénysejt saját maga védekezik, addig az állati szervezetben védekezésre specializálódott sejtek (makrofágok, granulociták, ölősejtek, limfociták) végzik az őrjáratot, szervezik meg és veszik fel a harcot (Erdei, 2003). Ezeknek a sejteknek a kialakulása (csontvelő, csecsemőmirigy, nyirokcsomó) sokszor távol esik a fertőzés helyétől, és a helyszínre a vér- és nyirokfolyadék áramlása útján jutnak el. Mivel a növények esetében ilyen cirkulációs rendszer nincsen, minden egyes szöveti sejt önmaga végzi el mind az általános, mind a specifikus (hiperszenzitív) védekezést. Ezért amikor összehasonlítjuk például a növényi általános védekezést az állati természetes, veleszületett immunitással, a megtámadott növénysejtet a fehérvérsejtekhez tartozó falósejtekkel (makrofágok, granulociták) kell összemérnünk, nem pedig egy állati szöveti sejttel (Klement, 2004) (1. ábra).
A növényekben a véráramhoz hasonló cirkulációs rendszer hiánya tette lehetetlenné a gerincesekre jellemző humorális védekezési rendszerek (ellenanyagok, citokinek által mediált folymatok) kialakulását.
A növényi általános (eredendő) rezisztencia és az állati természetes (veleszületett) immunitás hasonlósága
Az ember és más gerincesek esetében csak tíz év óta ismert, hogy a betolakodó mikrobát először a természetes, veleszületett immunrendszer ismeri fel, és azonnal támadást indít ellene. Szintén csak az elmúlt évek kutatásai bizonyították, hogy a növényekben is hasonló folyamat játszódik le (Klement et al., 2003). Még meglepőbb - mint ahogyan azt később látjuk -, hogy a növények általános, nem specifikus rendszere leginkább az állatok természetes, veleszületett immunitására hasonlít.
A veleszületett vagy általános immunitás, bár külsőleg gyakorlatilag tünetmentesen játszódik le, mikroszkóposan azonban mind a baktériumokra vadászó fehérvérsejtekben (neutrofil granulociták), mind a növények védekező sejtjeiben a fertőzés hatására különös hasonlóságot lehet felfedezni.
A növénysejt baktériummal érintkező részén, lokálisan, a citoplazmából vezikulumok, kis gömböcskék, granulumok alakulnak ki. Az ezekből kiszabaduló anyagból fenolokban és kallózban gazdag papillák alakulnak ki, melyek a a sejtmembrán és a szilárd sejtfal közötti térben helyezkednek el. Egyidejűleg sejtfalvastagodás és a baktériumot fokozatosan körülölelő hártya alakul ki, amely a baktériumsejt immobilizálásához és pusztulásához vezet (Ott és mts. 1997) (2. ábra).
Hasonló jelenséget láthatunk a fagocitózis esetében is. Pl. a fagoszóma kialakulásakor membrán vezikulumok, granulumok koncentrálódnak a baktériummal érintkező sejtmembrán részeknél, és ugyanakkor a baktériumsejtet (sejteket) körülölelő hártya képződik (2. ábra B).
Mindkét esetben jellegzetes, hogy a bezárt mikroorganizmus a külvilágtól leválasztott térben koncentráltan találkozik a számára toxikus anyagokkal. A granulumok fontos eleme a NADPH-oxidáz enzim, amely oxigén felhasználásával szuperoxid-anionokat (O2·-) állít elő. Ezek más enzimek segítségével erősen toxikus, reaktív gyökökké (ROS) alakulnak, amelynek egyik fontos tagja a hidrogénperoxid (H2O2). Ezen mérgező anyagok jelenléte a granulumokban, a sejtfalban és később az elpusztult baktériumsejtekben festéssel kimutatható. Ezek a jelenségek közös molekuláris folyamatokra utalnak.
Mind a növények, mind az állatok először a mikroorganizmus nem specifikus, ún. konzervatív, (az evolúció során is megőrződött) felületi elemeit ismerik fel. Számos ilyen felületi molekulát írtak le, de talán legismertebbek a baktériumsejtfalat alkotó lipopoliszacharid (LPS), az LPS-fehérje komplex vagy a baktériumcsilló fehérje monomerje, a flagellin.
A flagellin és az LPS ideális elicitorok mind a növényi, mind az állati általános vagy természetes immunrendszer beindítására, mert eukarióta sejtben nem fordulnak elő. Így ezeket mind a növényi, mind a védekezésre specializálódott állati sejtek (makrofágok, granulociták), mint idegen anyagot, felszíni receptoraik segítségével érzékelik és ismerik fel. A mikroba-felszíni elicitorokat összefoglaló néven PAMP-nak (pathogen associated molecular patterns) nevezik. Megjegyezzük, hogy a "pathogen associated" megjelölés nem helyes, mert ilyen felületi, konzervatív molekulákat mind a patogének, mind a szaprofiták egyaránt hordoznak. Éppen ezért általános elicitoroknak is hívjuk őket.
A növényi sejtek, a makrofágok és a granulociták felületén lévő jelfogó receptorok is sok hasonlóságot mutatnak. Ilyen felismerő rendszert először az ecetmuslicákban (Drosophila) vizsgáltak. A receptort kódoló gént Toll-nak nevezték el, majd kiderült, hogy a Toll típusú receptorok TLR (Toll-like receptor) evolúciósan megőrződtek, homológjaik megtalálhatók növényekben és emlősökben egyaránt (3. ábra) (Aderem and Ulevitch, 2000). Ezek a TLR típusú receptorok egy sejtfelszíni és egy intracelluláris domainból állnak. A sejtfelszíni LRR (leucin rich repeat) rész növényekben, rovarokban, állatokban egyaránt felismerhető. Az intracelluláris rész eltérő kinázokból áll (Shiu and Bleecker, 2001).
A sejtfelszíni receptorok azonnal üzenetet küldenek a sejtmag felé az idegen mikroorganizmus jelenlétéről. Ez a sejten belüli üzenetküldés a mitogén aktivált protein kinázok (MAPK) kaszkádjain keresztül történik (4. ábra). A kaszkád egymást foszforiláló kinázokból áll (MAPKKK, MAPKK és MAPK), amelyek akkor lépnek működésbe, ha a receptor felől jelzés érkezik (Chang és Karin, 2001). A MAPK kaszkádok szintén evolúciósan megőrzött modulok, amelyek növényben, rovarban, emlősökben alapvető szabályozó szerepet töltenek be, és a jelátvitel során hidat képeznek receptortól a célgénekig (Tena et al., 2001). Eddig két növényi MAPK kaszkádot derítettek fel részletesen, egyet a dohányban és egyet az Arabidopsisban. E teljes útvonal feltérképezésével lehetőség nyílt a Drosophilában és az emlősökben zajló folyamatokkal való összehasonlítására, amelyekkel meglepő párhuzam mutatkozik (Asai et. al. 2002) (3. ábra).
Figyelemreméltó, hogy a patogének (kompatibilis kapcsolatban) olyan támadó mechanizmust fejlesztettek ki, amely képes meggátolni az általános, vagy veleszületett védekezési rendszert. Így pl. lehetetlenné teszik a növényekben az általános rezisztenciára jellemző papillaképződést, a H2O2 termelést, a sejtfalvastagodást és baktériumbecsomagolást. Mindezzel viszont lehetővé válik a virulens patogén szaporodása (Bozsó, 2001). Ezt pl. a baktériumsejtben termelt szuppresszor fehérjéknek tulajdonítják, amelyek képesek elnyomni a MAPK kaszkád alatti folyamatot (Cornelis, 2002). Ilyen szuppresszor fehérjéket csak a kompatibilis kapcsolatban lévő patogén baktériumok állítanak elő, amelyek az ún. III. típusú szekréciós rendszer segítségével jutnak a megtámadott növénysejtbe. Feltételezik, hogy ezek a szuppresszor fehérjék bizonyos vir gének kifejeződésének a termékei. Állat-patogének pl. Yersinia enterocolitica szupresszor fehérjéi (YopE, YopH, YopT és a YopO) hasonló mechanizmus szerint gátolják a makrofágokban a fagocitózist, az oxidatív robbanást azáltal, hogy megakadályozzák a MAP kináz aktiválását a szignál folyamatban. Úgy tűnik tehát, hogy a növény patogén baktériumok hatására termelődő effektorok hasonló módon szuppresszálják a növényben az általános rezisztenciát, mint ahogyan az a fagocitózisra képes állati sejtekben történik.
A növényi specifikus (hiperszenzitív) rezisztencia és az állati adaptív (szerzett) immunitás összehasonlítása
A törzsfejlődés során a különböző fejlődési vonalra lépett növényekre és gerincesekre specifikus kórokozók fejlődtek ki, melyekkel szemben az evolúció későbbi szakaszában már specifikus védekező rendszert kellett kiépíteni. Ezt a specificitást a törzsfejlődés magasabb fokán kialakult növény- és állatfajokon belüli új fajták megjelenése tovább fokozta. Ennek köszönhető, hogy az általános rezisztencia egymagában már nem nyújtott megfelelő védelmet, és így a növényvilágra és az állatvilágra jellemző különböző specifikus védekezési mechanizmusok fejlődtek ki. Bár ezek az új védekezési mechanizmusok már külön utakra léptek, és nehezen összehasonlíthatók, mégis egy-egy közös vonás ezekben is fellelhető.
A baktériumok elleni specifikus védekezési mechanizmus a növényvilágban az ún. hiperszenzitív rezisztencia (Klement et al., 1964), a gerinceseknél pedig az adaptív immunitás.
A specificitást a növényeknél a fajra és fajtára jellemző specifikus R (rezisztencia) intracelluláris receptor gének, a gerinceseknél pedig a specifikus, antigént felismerő T-és B-limfociták biztosítják. A limfociták felszíni receptorainak variabilitását számos génszegmentum kódolja, amelyek véletlenszerűen átrendeződnek (rekombinálódnak) a limfociták egyedfejlődése során. Növények esetében az R-gének a növénykórokozók Avr géntermékeivel, míg a gerinceseknél a limfocitareceptorok a makrofágok és dendritikus sejtek segítségével szállított és bemutatott, mikroorganizmus-eredetű finomstruktúrákkal, az ún. antigénekkel lépnek kapcsolatba. Ezeknek a specifikus receptoroknak az aktiválódása vezet el a nagymértékű, fajlagosságot biztosító hiperszenzitív, illetve adaptív immunválasz kialakulásához. A növények specifikus, hiperszenzitív védekezési rendszere jelentősen eltér az állati immunrendszertől. Azonban az R-gének specificitásának kialakulása emlékeztet a limfociták nagyfokú sokféleségének létrejöttére. A növényi R gének gyakran génegyüttesekben, ún. klaszterekben egymáshoz közel helyezkednek el. Ez felveti a hasonló gének közötti gyakori rekombináció lehetőségét és ezáltal új öröklődő variációk (új fajtarezisztencia) létrejöttének esélyét (Boller és Keen, 1999). Gerinceseknél hasonló lehetőség van az adaptív immunrendszerben fontos szerepet játszó limfociták génjeinek szomatikus átrendeződésére egy adott egyeden belül, ami által végtelen számú patogén felismerése lehetséges, de ez nem öröklődik. A limfocitákra jellemző, hogy aktiválódásukat követően nem csak effektorsejtek, hanem hosszú életű memóriasejtek is kifejlődnek. Ez a mechanizmus biztosítja azt, hogy a kórokozó ismételt támadása esetén rövid idő alatt nagyon hatékony immunválasz alakul ki. Az immunológiai memória tehát az egyed élete alatt, a limfociták aktiválását követően fejlődik ki, és ezért nem öröklődik a következő nemzedékre. - Bár a növényeknél ilyen értelmű memória nincsen, egy bizonyos fajta memóriát az új kórokozó biotípusok megjelenésére válaszként kialakuló új, specifikus R gének biztosítanak, amelyek viszont nemzedékről nemzedékre öröklődnek.
További közös vonást találunk az állat- és növénykórokozó baktériumok fertőzési technikájában is: nevezetesen a kórokozók géntermékeinek bejuttatását a gazdasejtbe. Erre a célra a növényi és egyes állati kórokozó baktériumok ún. III. típusú szekréciós rendszert fejlesztettek ki, mely a kórokozók közös ősből származó kialakulására enged következtetni. A növényeknél a hiperszenzitív nekrózis kialakulása emlékeztet a fehérvérsejtekhez tartozó neutrofil granulocitákban lejátszódó folyamathoz, ahol többek között a felhalmozódó ROS hatására nemcsak a kórokozó, de a fehérvérsejt maga is elpusztul. Amikor ez a folyamat nagy mennyiségű fehérvérsejt feláldozásával jár, akkor az gennyképződés formájában jelentkezik. Ezért az indukált növényi szövetelhalást a gennyképződés folyamatával analógnak tekinthetjük.
Az általános és hiperszenzitív védekezés lehetséges kapcsolódása
A gerinceseknél a természetes és szerzett immunitás egymásra épülése bizonyított. A természetes immunitásban részt vevő különböző sejttípusok kettős feladatot látnak el. Egyrészt felismerik az idegen mikrobát, és azt közvetlenül is károsítják, másrészt közvetítik az idegenről szóló információt az adaptív immunrendszer számára.
A növényeknél ilyen szoros kapcsolatot az általános és hiperszenzitív védekezési rendszerben annak ellenére sem találunk, hogy egyazon növénysejten belül mindkét mechanizmus szinte párhuzamosan fejlődik ki. Előfordul, hogy az általános védelem megelőzi a hiperszenzitívet, amikor is az előbbi meggátolja a második kifejlődését. Ilyen eset akkor áll elő, amikor az általános rezisztenciához tartozó korai indukált rezisztencia (EIR) olyan gyorsan fejlődik ki, hogy az leblokkolja a HR indukcióját. Másrészt viszont a hiperszenzitív védekezéssel kapcsolatos III. típusú szekréciós rendszer kompatibilitás esetén a szuppresszor fehérjék növénysejtbe juttatásával az általános rezisztencia gátlását eredményezi. Itt is érzékelhetjük a két rendszer kapcsolódását.
A két védekezési rendszer szoros együttműködését bizonyítja laboratóriumunkban Bozsó Zoltán és munkatársainak legutóbbi genomikai vizsgálata is. A Medicago truncatula fertőzésekor figyelték az általános (EIR) és a hiperszenzitív védekezési reakciók alatt aktiválódott és represszálódott gének számát. A fertőzést követő 6. órában a HR és az EIR alatt 16 000 génből 2771 (16 %) gén aktivitása változott meg szignifikánsan. A 2774 génből 698 (25 %) gén közös volt, vagyis mindkét védekezési mechanizmusban részt vettek. A közös gének nagy száma szintén a két mechanizmus együttműködésére utal.
Evolúció: a túlélés stratégiája
Minden élőlény kidolgozta a túlélés stratégiáját. Ezt tapasztaljuk a fertőzésen átesett növények és állatok populációja esetében is, ahol a populáció egyes egyedei mutáció segítségével túlélik a fertőzést, alapot adva az új, rezisztens, betegségnek ellenálló populáció kialakulására. Azonban a kórokozók is kifejlesztették a saját túlélési stratégiájukat. Ennek eredménye a gazda és a patogén koevolúciója, vagyis a cél a faj eredményesebb fennmaradása, legyen az növény-, állat- vagy patogénfaj. Ezt az evolúciós folyamatot láthatjuk a baktérium-növény kapcsolatában is az 5. ábrában felvázolva.
Erdei Anna előadásában szép példákat találunk a természetes és adaptív védekezési rendszerek törzsfejlődés alatti kialakulására. Hasonlóan érdekes adatokat ismerhetünk meg Andó István dolgozatából, a Drosophyla különböző fejlődési alakjaiban működő mechanizmusok kapcsán. A növényekben a törzsfejlődés alatti folyamatokról pontos információink nincsenek, csak bizonyos következtetésekre vagyunk utalva.
Feltételezhető, hogy a mikroorganizmusok az élőlényekben táplálékot keresve igyekeztek megtelepedni. Ennek megakadályozására fejlesztették ki az élőlények az általános védekezési mechanizmust. Ennek a harcnak egyensúlyi állapotát jelzik az ún. szimbionta baktériumok (pl. Rhizobium), amelyek már kölcsönös együttélésre rendezkedtek be. Bozsó és munkatársainak vizsgálatai azt mutatták, hogy pl. Rhizobium meliloti nitrogénkötő baktérium az általános rezisztenciát képviselő korai indukált rezisztenciát a Medicago növényben nem indukálja vagy közömbösíti. Talán ennek köszönhető, hogy az együttélés fennmaradhatott.
Az evolúció következő szakaszában, amikor a vizekben, talajban vagy a növények felületén nagy számban élő Pseudomonas fluorescens szaprofiton baktériumhoz rokonságilag nagyon közel álló patogén Pseudomonas patotípusok már kialakultak, akkor az általános, nem specifikus immunrendszer önmagában már nem volt elegendő ezek fékentartására. Erre az átmeneti állapotra példa lehet az opportunista, számos növényt fertőző (polyvirulens) patogén Pseudomonas syringae pv. syringae, amely a növény általános immunrendszerének meggyengülése esetén (például lehűlések alkalmával) képes tömegesen felszaporodni és súlyos betegséget okozni (Besenyei et al., 2003).
A baktériumpatogének saját gazdanövényükben az általános védekezési mechanizmust - annak ellenére, hogy indukálják, mégis - "átlépik" (vagy közömbösítik), és betegséget okoznak (Bozsó et al., 2001). Feltételezhetően ezek ellen építette ki a növény a kórokozóra nézve specifikus hiperszenzitív védekezési rendszert, ahol a növény R receptor génje vagy terméke a kórokozó avr gén termékét felismeri és a HR-t beindítja. Azonban egyes túlélő baktériumok között olyan új mutánsok, kórokozó rasszok vagy biotípusok alakulhatnak ki, amelyek már új avr génkészlettel rendelkeznek. Ezek a mutánsok a korábban rezisztens növényben már nem indukálják és így kikerülik a hiperszenzitív választ, és ezért betegséget okoznak. Ugyanakkor a növények is - a túlélési stratégiát követve - új fajtákat hoznak létre, amelyek R génjei már képesek felismerni az új kórokozó biotípust, és így betegség-ellenállók lesznek. Ennek a folyamatnak köszönhető, hogy korábban rezisztens fajták 8-10 év alatt fogékonyakká válnak. Ebből a megfigyelésből kiindulva a növénynemesítőknek olyan betegség-ellenállósági stratégiát kell kiépíteniük, amelynek az alapja nem a rövid életű hiperszenzitív rezisztencia, hanem sokkal inkább az általános, nem specifikus védekezési rendszer felerősítése, ami már széles skálájú rezisztenciát biztosíthat.
A patogének nem csak új avr géneket hordozó biotípusokat vagy rasszokat hoztak létre, hanem más mechanizmusokat is alkalmaztak létük fenntartása érdekében. Ilyen a növénykórokozó baktériumokra jellemző burok, amely több funkciója mellett fontos feladatot lát el a növény-kórokozó kapcsolatban. Például fizikailag meggátolja a növényi és a baktériumsejtfal közötti közvetlen érintkezést, és ezzel elkerüli a felismerést és ezáltal a növény védekezési mechanizmusának beindítását (lásd Klement előző dolgozatát). Király Zoltán munkatársaival (1997) kimutatta, hogy ez az extracelluláris burok a növény védekezésekor keletkezett szabad gyökök káros hatásától is megvédi a baktériumsejteket, ami egyben lehetővé teszi a kórokozó tömeges szaporodását is. A burokképződésre az állatpatogének között is számos példát találhatunk.
Általános következtetések
Mint láthattuk, a növények és állatok védekezési rendszerében számos közös vonás fedezhető fel (1. ábra). Ez különösen érvényes az általános, illetve a természetes immunrendszerre, ami feltételezhetően közös eredetű, és amit mind a növények, mind az állatok az evolúció folyamán megőriztek. Ez a közös vonás különösen tetten érhető molekuláris szinten, ahol a mikroba felületi molekuláit, ún. elicitorokat (PAMP) a hasonló felépítésű, (LRR-típusú) receptorkinázok ismerik fel. A MAPK kaszkád szintén univerzális jelátviteli mechanizmus, amely összeköti a különböző receptorokat a sejtbeli és sejtmagbeli válaszmechanizmusokkal a növényben, Drosophilában és emlősökben. Az oxidatív robbanás a növényeknél és állatoknál is fontos faktor a védekezésben.
A védekezési mechanizmusok a fejlődés magasabb fokán külön utakra léptek, létrehozva a növényeknél a specifikus, hiperszenzitív, az állatoknál az adaptív, fajlagos védekezési rendszereket. Ezek, bár jellemzően különbözőek, mégis bizonyos elemeiben közös vonásokat hordoznak magukban.
Jellemző eltérés a humorális védekező-rendszer hiánya a növényi védekezési rendszerben. Ennek feltehetően elsődleges oka a gerincesekben meglévő cirkulációs rendszerek (vér, nyirok) hiánya a növényeknél.
Úgy érzem, hogy elcsodálkozhatunk azon, hogy a növényi és állati immunrendszernek milyen sok közös vonása van. Azonban az evolúciót ismerve inkább azon kellene elcsodálkoznunk, ha ez nem így volna. Valóban érdemes elgondolkoznunk viszont azon, hogy eddig nem foglalkoztunk eléggé ezen rendszerek közös eredetével, és hogy a növényi, ill. állati szervezetekkel foglalkozó immunológusok kölcsönös érdeklődése csupán az utóbbi években fejlődött ki. Ez a közös érdeklődés mindkét oldalnak új ötleteket adhat. Mi, növény-patológusok nem vagyunk abban a szerencsés helyzetben, hogy több Nobel-díjassal is dicsekedhessünk, de talán szerény hozzájárulásunk lehetővé teszi különösen az általános vagy természetes immunitás gyökereinek jobb megértését.
Kulcsszavak: általános rezisztencia, veleszületett immunitás, hiperszenzitív rezisztencia, adaptív immunitás, molekuláris patológia
1. ábra * Védekezés az élővilágban, az állati és növényi védekezés közös és
eltérő tulajdonságai
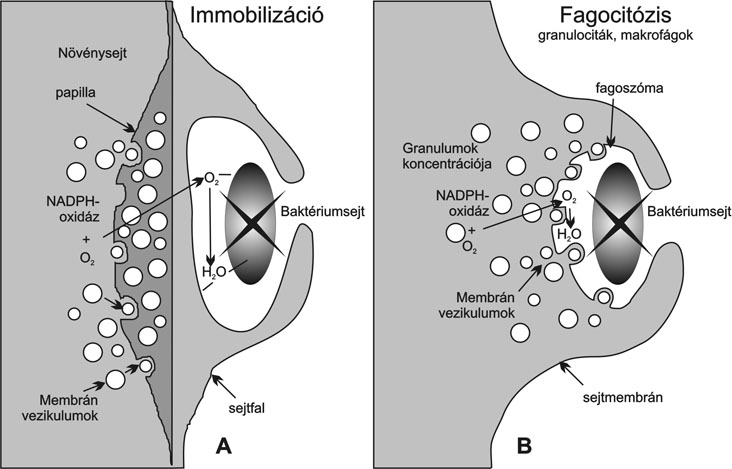 2. ábra * Az immunizáció és a fagocitózis hasonló mechanizmusa a növényi
sejtben, és a granulocitákban illetve makrofágokban
2. ábra * Az immunizáció és a fagocitózis hasonló mechanizmusa a növényi
sejtben, és a granulocitákban illetve makrofágokban
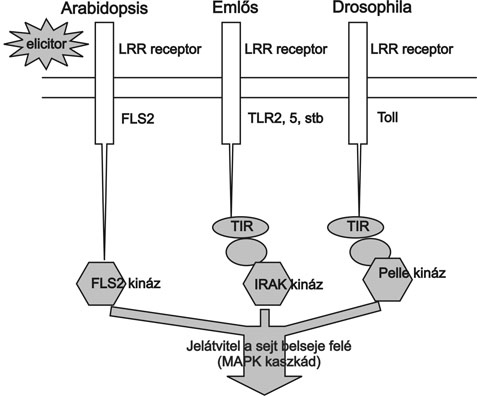 3. ábra * Érzékelés és jelátvitel modellje Arabidopsis, emlős és Drosophila
veleszületett immunrendszerében (Asai 2002 nyomán)
3. ábra * Érzékelés és jelátvitel modellje Arabidopsis, emlős és Drosophila
veleszületett immunrendszerében (Asai 2002 nyomán)
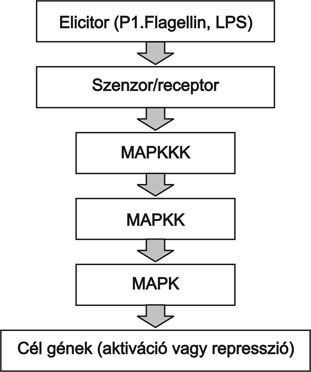 4. ábra * A MAPK kaszkádok működésének sematikus diagramja (Tena, 2001 nyomán)
4. ábra * A MAPK kaszkádok működésének sematikus diagramja (Tena, 2001 nyomán)
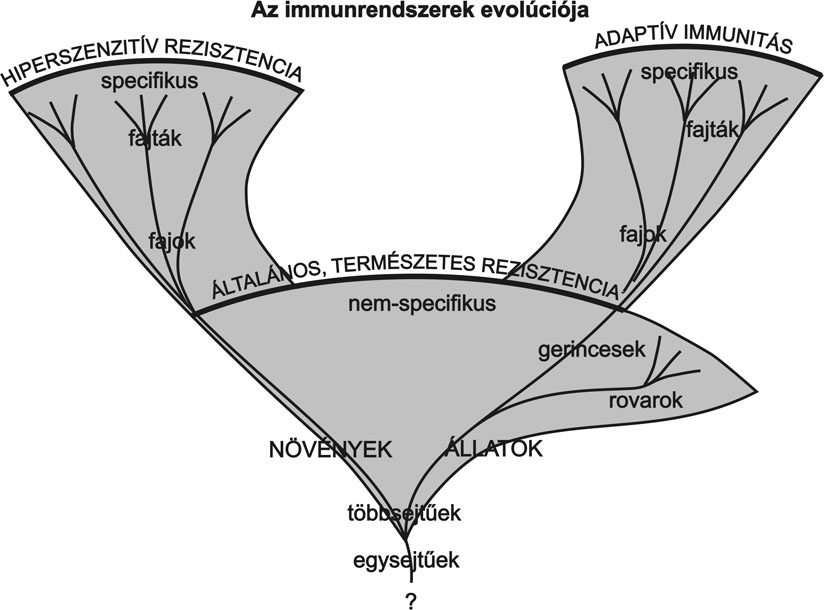 5. ábra * Az immunrendszerek evolúciója
5. ábra * Az immunrendszerek evolúciója
Szószedet
Antigén: Mindazon sejtek, molekulák, melyeket az emlősök érett immunrendszere felismer.
Antitest (ellenanyag, immunglobulin): Antigénekre specifikus receptorok, melyek a B limfociták membránján fordulnak elő, onnan leválva, a keringésbe kerülve felismerik a kórokozót.
Apoptózis: A sejt programozott halála, melynek során a sejt és a DNS feldarabolódik, végül maradványait (emlősöknél) fagocita sejtek kebelezik be. Növényeknél fontos szerepe van a hiperszenzitív reakcióban.
Avr-gének: A kórokozók speciális avirulencia génjei, melyek termékei a növényi sejtbe jutnak, és itt az R gének termékeivel kölcsönhatásba lépnek.
Elicitor: Olyan, mikrobák által termelt molekulák, amelyek a növényben válaszreakció elindítására képesek.
EPS (extracelluláris poliszacharid): A legtöbb növényi kórokozó baktérium cukor jelenlétében EPS burkot képez maga körül, mely víz megkötésével segíti a betegség kialakítását.
Fagocitózis: Állatokban jellemző folyamat, melynek során egy sejt szilárd részecskét (akár másik sejtet) kebelez be.
Flagellin: A baktériumok ostorát felépítő fehérje építőköve (monomerje).
Gén-génnel szembeni rendszer: Elmélet, mely szerint egy kórokozó és egy növény kapcsolata akkor lesz rezisztens, ha a kórokozó rendelkezik olyan avirulencia génnel, mellyel szemben a növényben jelen van egy specifikus rezisztencia gén.
H2O2 (hidrogénperoxid): A reaktív oxigénformákhoz tartozik, az élő szervezetek stressz esetén termelik. A kórokozókat hatékonyan pusztítja, de magát a termelő sejtet is károsíthatja, pl. a HR esetében.
HR (hiperszenzitív reakció): A patogén mikroorganizmus által a nem-gazda növényen és növényfajtán kiváltott válaszreakció. Gén-gén (R és avr) kölcsönhatás eredményeképpen kialakuló sejtelhalás.
Hrp génegyüttes: A baktériumok III. típusú szekréciós rendszerét kódoló génegyüttes, melynek segítségével a kórokozó a patogenitásban részt vevő molekulákat juttathat a megtámadott sejtbe.
Limfociták: Az emlősök adaptív immunrendszerének kulcsfontosságú sejtjei, melyek biztosítják a rendkívül specifikus immunválaszt, és az immunológiai memóriát.
LPS (lipopoliszacharid): A Gram-negatív baktériumok jellegzetes sejtfal alkotórésze.
LRR (leucine rich repeat): Leucinban gazdag ismétlődés. Egység a fehérjéken (általában receptorokon) belül, amely képes kölcsönhatásba lépni más fehérjékkel.
MAPK (mitogén aktivált protein kinázok): Olyan, jelátvitelben résztvevő kinázok (más fehérjéket foszforiláló enzimek), melyek akkor aktiválódnak, ha egy másik enzim őket is foszforilálja. Ezáltal foszforilációs sorozatot (kaszkádot) alkothatnak.
NBS (Nucleotide binding domain): Nukleotid kötő motívum.
Neutrofil granulociták: a fehérvérsejtek (leukociták) egy csoportja, melyek fő feladata a baktériumsejtek elleni védekezésben az idegen anyagok elpusztítása fagocitózis által.
Oxidatív robbanás: Reaktív oxigénformák (ROS) és származékaik hirtelen felszabadulása. Növényi sejtek, és emlősök fagocita sejtjei (neutrofil granulociták) is képesek előidézésére.
PAMP (pathogen associated molecular patterns): A mikroorganizmusokon általánosan előforduló molekulamintázatok, melyek indukálják az általános, nem specifikus védekezési mechanizmusokat.
Receptor: Jelfogó molekula, mely ha kapcsolatba lép az érzékelendő molekulával, jelet küld a sejt belseje felé. Így informálja a sejtet a külvilág változásairól, amire a sejt belsejéből válaszreakció érkezik.
R-gének: A gazdanövények specifikus rezisztenciagénjei.
RLK (Receptor-like kinase): receptorszerű kináz. Olyan molekula, amely rendelkezik felismerő (receptor), és foszforiláló (kináz) résszel.
ROS (Reaktív oxigén formák): Az oxigénből származtatható vegyületek, melyek könnyen lépnek reakcióba más molekulákkal, így az élő szervezetben károsíthatják például a DNS-t, RNS-t, fehérjéket.
TIR domén (Toll/Interleukin-1 Receptor): A Toll-, a Toll-szerű-, és a humán Interleukin-1 receptoron megtalálható jellegzetes motívum.
TLR (Toll like receptors): Toll típusú receptorok. A Drosophila Toll nevű receptorával azonos típusú fehérjék.
Toll receptor: Drosophila membránt átívelő receptora, mely a sejten kívül egy leucingazdag ismétlődést (LRR), a sejten belül egy TIR motívumot tartalmaz. Az embrióban az egyedfejlődésben van szerepe, a kifejlett állatban a veleszületett immunrendszer fontos eleme.
Transzgenikus növény: Olyan növény, melybe géntechnológiai eljárásokkal egy meghatározott szerepű gént ültettek be, akár egy hasonló növényből, akár bármely más szervezetből.
Vakuolum: A növénysejtek belsejében található zsákszerű szervecske. A sejt itt raktároz később hasznosítandó anyagokat, valamint mérgező salakanyagokat is.
Irodalom
Aderem, A., Ulevitch, J. (2000). Toll-Like Receptors in the Induction of the Innate Immune Response. Nature. 406, 782-787
Asai. T., Tena, G., Plotnikova, Joulia, Willmann, M. R., Chiu, W-L., Gomez-Gomez, L., Boller, T., Ausubal, F. M., Sheen, J. (2002). Map Kinase Signalling Cascade In Arabidopsis Innate Immunity. Nature. 415, 977-983
Besenyei, E., Bozsó, Z., Ott, Pg., Czelleng, A., Szatmári, Á., Varga, G., és Klement, Z. (2003). A növényi védekezési mechanizmusok elmaradása és az alacsony hőmérséklet szerepe az opportunista patogén, Pseudomonas syringae pv. syringae baktérium kolonizációjára. 49. Növényvédelmi Tudományos Napok, Budapest, Február 25-26. P. 87
Boller, T., Keen, N. T. (1999). Resistance Genes and the Perception and Transduction of Elicitor Signals in Host-Pathogen Interactions. In: Slusarenko, A., Fraser, R. S. S., Van Loon, L. C. (Eds) Mechanisms of Resistance to Plant Diseases. 189-229. Kluwer Academic Publishers, the Netherlands
Bozsó, Z., Ott, P. G., Kecskés, M. L., Czelleng, A., Klement, Z.. (2001). Non-Specific Peroxidase and H2O2 Associated Reactions of Tobacco Leaves After Infiltration with Different Hrp/Hrma Mutant of P. Syringae Pv. Syringae 61. In: Plant Pathogenic Bacteria. Proceedings of the 10th International Conference on Plant Pathogenic Bacteria. 195-198 Kluwer Academic Publishers, Dordrecht, the Netherlands
Chang, L., Karin, M. (2001). Mammalian Map Kinase Signalling Cascades. Nature. 410, 37-40
Cornelis, G. R. (2002). The Yersinia Ysc-Yop “Type III." Weaponry. Nat. Rev. Mol. Cell Biol. 742-752.
Erdei A. (2003). A természetes immunitás hatalma. Magyar Tudomány. 4, 422-429
Király, Z., El-Zahaby, H. M., Klement, Z. (1997). Role of Extracellular Polysaccharyde (EPS) Slime on Plant Pathogenic Bacteria in Protecting Cells to Reactive Oxygen Species. J. Phytopathology. 145, 59-68
Klement, Z. (2004). Önvédelem a Növényvilágban. Mindentudás Egyeteme 2., Kossuth Kiadó, Budapest (megjelenés alatt)
Klement, Z., Bozsó, Z., Kecskés, M. L., Besenyei, E., Czelleng, A., Ott, P. G. (2003). Local Early Induced Resistance of Plants as the First Line of Defence Against Bacteria. Pest Management Science. 59, 465-474
Klement, Z., Farkas, G. L., Lovrekovich, L. (1964). Hypersensitive Reaction Induced by Phytopathogenic Bacteria in the Tobacco Leaf. Phytopathology. 44, 474-477
Ott, P. G., Szabó, L., Balázs, E., Klement, Z. (1997). Submicroscopic Evidence of Bacterially Induced Resistance in Tobacco Leaves. Acta Phytopathologica et Entomologica Hungarica. 32(3-4), 265-280
Shiu, S-H., Bleecker, A. B. (2001). Receptor-Like Kinases from Arabidopsis Form a Monophyletic Gene Family Related to Animal Receptor Kinases. PNAS. 98(19), 10763-10768
Szatmári, Á. és Klement, Z. (2003). Hasonlóságok és különbözőségek a növény- és állatvilág immun-mechanizmusában. Növénytermelés. 52. 6. 703-712.
Tena, G., Asai. T., Chiu, W-L., Sheen, J. (2001). Plant Mitogen-Activated Protein Kinase Signaling Cascades. Current Opinion in Plant Biology. 4, 392-400