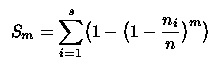
Fajközösségek egyféle számszerű jellemzésének céljából induljunk ki a közösségben előforduló fajok egyedszámainak n1, n2, ..., ns együtteséből. Tehát - mellőzve számos szóba jövő aggályt - hagyjuk figyelmen kívül az egyes gyakoriságok fajok szerinti hovatartozását. Természetesen azonnal felvetődik az a probléma, hogy a megfigyelt egyedszámok és a fajszám nagymértékben függhet a vizsgált fajközösség vagy az ebből származó minta nagyságától. A tárgyalás egyszerűsítése céljából azonban ezzel a kérdéssel és általában a mintavétel kérdésével nem foglalkozunk. Az intuitíve értelmezett faji diverzitás valamely mérőszámától elvárjuk, hogy értéke rögzített fajszám mellett növekedjék, amennyiben az egyedek fajok körében való eloszlási egyenletessége növekszik, vagyis ha a megoszlás koncentráltsága csökken. Függjön továbbá valamilyen módon a mérőszám értéke az előforduló fajok számától is. Ez utóbbi követelmény leegyszerűsödik, egyszersmind konkrétabbá válik, ha azt feltételezzük, hogy a teljesen egyenletes megoszláshoz tartozó indexérték növekedjék a fajszám növekedésével. A diverzitási index értékének kialakításában tehát két összetevő, az úgynevezett egyenletességi (evenness) komponens és a fajszám vagy fajgazdagsági (richness) komponens játszik szerepet. Ami itt a komponens kifejezést illeti, csupán intuitív megközelítésre utalhatunk, mert a használatos diverzitási indexek nem állíthatók elő az egyenletességi és fajbőségi komponens konkrét függvényeként, például összegként vagy szorzatként.
Diverzitási mérőszámként igen nagyszámú függvényt vezettek be a statisztikus ökológiában (lásd Izsák, 2001; Tóthmérész, 1997). Másrészt a közgazdaságtanban, bibliometriában, informetrikában használatos koncentráltsági indexek (Egghe, 1991; Frigyes, 2002) monoton fogyó függvényei (ellentettjük, reciprokuk stb.) egyenletességi mérőszámoknak tekinthetők, és rögzített fajszámot feltételezve diverzitási indexként is használhatók. A diverzitás mérésével foglalkozó sokezer közlemény és számos monográfia a diverzitási indexek számos statisztikai tulajdonságát tárta fel, és számos ökológiai tanulsággal szolgált. Mindez ma már a statisztikus ökológia integráns részét képezi. Hazánkban 2001-ben tartottak nemzetközi konferenciát a (faj)diverzitás és rokon területek témakörében. A konferencián kialakult általános vélemény szerint a fajdiverzitásra kizárólagos meghatározás nem adható, és az egyes diverzitási indexek a fajdiverzitásnak kisebb-nagyobb mértékben eltérő értelmezésén alapulnak. Így nem csoda, hogy a diverzitási szakirodalom napjainkban is folyó ütemes bővülésének ellenére a diverzitási indexek ökológiai alkalmazhatóságával szembeni szkepticizmus nem szűnik.
Visszatérve a fajgazdagsági komponenshez, azaz magához a fajszámhoz, ez a környezetvédelemben mindennaposan használt biodiverzitás mértékeként is azonosítható. Maga a biodiverzitás - a fajdiverzitástól eltérően - nem egyéb, mint a fajszámra vonatkozó, egyfajta minősítést is magában foglaló kifejezés.
Mint említettem, a szakirodalomban leírt nagyszámú diverzitási index adott fajközösség diverzitását más és más oldalról mutatja be. Olyan eset is előfordulhat, hogy valamely A és B fajközösség esetén egy diverzitási index A diverzitását mutatja nagyobbnak, egy másik index B diverzitását (Patil - Taillie, 1979; Tóthmérész, 1997). Ez azonban viszonylag ritka eset. Általános a tapasztalat, hogy nagyszámú állat- vagy növényközösség kétféle indexszel mért diverzitása jelentős pozitív korrelációt mutat. Ez egyrészt megnyugtató körülmény, hiszen arra utal, hogy az egyik index a másik indexhez hasonlóan tudósít a fajdiverzitási viszonyokról. Ugyanakkor többféle diverzitási index alkalmazását éppen az indokolhatja, hogy azok a diverzitási viszonyokat más-más oldalról világítják meg. Az említett pozitív korreláció legfőbb oka az indexek különösen nagyfokú érzékenysége a legnagyobb fajgyakoriság parciális (a többi gyakoriság változatlanul hagyása melletti) növelésére. Ha például parciálisan 5 %-kal növeljük egy fajközösségben a legnagyobb "2000" gyakoriságot, akkor az indexérték változásának (csökkenésének!) abszolút értéke sokszorosa annak, mint amit az ugyanezen fajközösségben észlelt "20" egyedszám 5 %-os növelése előidéz (utóbbi változás mindig pozitív). Míg tehát a diverzitási indexek (és hasonlóképpen a koncentráltsági indexek) függvényként elvileg nagyszámú gyakorisági értékre támaszkodnak, addig a gyakorlatban csupán a néhány legnagyobb gyakoriság játszik lényeges szerepet az indexérték kialakításában. A kérdéskör részletesebb elemzése az ún. indexérzékenységi vizsgálatok körébe tartozik. Ezek módszertani alapjainak megteremtése kezdetben nem vezetett eredményre. Megfelelő módszer bevezetésével az 1990-es évektől kezdődően vált lehetővé az indexérzékenység matematikai elemzése (Izsák, 1996). Az 1a és 1b ábrán a Shannon-féle diverzitási indexre vonatkozó deltani érzékenységi értékeket mutatjuk be adott gyakoriságegyüttes ni (illetve a szemléltetés érdekében a logni értékeknek) függvényében. Az 1a ábrán látható érzékenységi grafikon jól mutatja, hogy a legnagyobb gyakoriság növelése a diverzitási index különösen nagymértékű változását (éspedig csökkenését) eredményezi. A legnagyobb gyakoriság és a hozzá tartozó érzékenység elhagyásával adódó 1b ábra viszont jól mutatja, hogy az érzékenységi grafikon pozitív értéktartománybeli szakasz után maximumot ér el, majd mérsékelt, végül meredek csökkenést mutat, miközben negatív értéktartományba lép. Eközben a grafikon metszi a gyakorisági tengelyt. A metszésponthoz közeli gyakoriságot (közelítően) neutrális gyakoriságnak nevezhetjük, amennyiben ezen gyakoriság csekély növelése a diverzitás értékét gyakorlatilag nem befolyásolja.
A grafikon alapján azt is megállapíthatjuk, hogy az érzékenység előjelének tekintetében a gyakoriságok két tartományba sorolhatók: egyik részük csekély növelése a diverzitási index értékét növeli, másik részük esetében ugyanilyen esetben az indexérték csökken. Mindig ez utóbbi tartományhoz tartozik a legnagyobb, domináns gyakoriság. Így - legalábbis adott diverzitási indexszel kapcsolatban - a neutrális gyakoriságnál nagyobb gyakoriságokat mondhatjuk domináns gyakoriságoknak (Izsák - Papp, 2002).
Kapható persze az első ábrához hasonló grafikon úgy is, hogy az egyes gyakoriságokat parciálisan, csekély és azonos arányban növeljük, és újraszámolás után regisztráljuk a diverzitás értékének megváltozását. Kis gyakoriságok esetében azonban tört gyakorisági értékek adódhatnak, ami máris módszertani nehézséget jelent. Például ez a probléma, de összetettebb kérdések megválaszolásának igénye is indokolta az említett, távolabbra mutató vizsgálati módszer kidolgozását. Ilyen kérdéssel foglalkozunk röviden az alábbiakban.
Ökológiai diverzitásvizsgálatokban gyakran használják az Sm diverzitási indexcsalád tagjait. E paraméteres indexcsalád képlete:
ahol n a mintaméret, azaz ni/n az i faj relatív gyakorisága. Amennyiben az n1, n2, ...., ns gyakoriságokkal rendelkező sokaságból visszatevéses eljárással m számú egyedet véletlenszerűen kiválasztunk, akkor az m elemű mintában reprezentált fajok számának várható értéke Sm. Felvethető a kérdés, hogyan befolyásolja az index-érzékenység pozitív tartományba eső lokális maximum helyét, valamint a neutrális gyakoriság értékét az m paraméter növelése. Megmutatható, hogy az m paraméter növelésekor mind a maximumhelynek megfelelő gyakoriság, mind a neutrális gyakoriság egyre kisebb lesz (Izsák, 1996). Ez a megállapítás hasznosítható az Sm indexcsalád általános statisztikai tulajdonságainak elemzésekor. Kívánatos volna hasonló vizsgálat végzése a Rényi-féle általánosított entrópiával kapcsolatban is.
A kvadratikus entrópiaindex
Első pillantásra kikezdhetetlennek tűnik a diverzitási indexekkel szembeni azon követelmény, hogy az index rögzített fajszám mellett egyenletes megoszlás esetében felvegye maximumát. Más azonban a helyzet akkor, ha a fajok közti eltérés különböző mértékét is figyelembe vesszük. A legegyszerűbb feltételezés természetesen az, hogy két egyed faj szerinti különbözősége 0 vagy 1 aszerint, hogy azonos vagy különböző fajhoz tartoznak. Ebben az esetben két, visszatevéses módon és találomra kiválasztott egyed különbözőségének várható értéke a fentiek szerint S2-nek az n1, n2 , ..., ns gyakoriságegyütteshez tartozó értéke, nevezetesen 2-Summa(ni/n)2. Ám ha az i és j faj különbözősége valamely általános dij érték és Delta a(dij), (i = 1, 2, ..., s, j = 1, 2, ..., s) mátrix, akkor a különbözőség várható értéke az r =(n1/n, n2/n,...ns/n) vektorjelöléssel a Q=r'Deltar kvadratikus entrópia. Ezt a diverzitási indexként alkalmazható mérőszámot jóval a klasszikus diverzitási indexek bevezetése után írták le a biometriai szakirodalomban (Rao, 1982). Az, hogy alkalmazásával a fajok közti különbségek eltérő mértékét is figyelembe vehetjük, új és egyelőre kiaknázatlan lehetőséget teremt a diverzitási (és koncentráltsági!) elemzésekben. Érdekességként említhető, hogy az egyedek faji különbözősége egyetlen faj egyedeit tekintve sem feltétlenül 0, mert egyetlen faj egyedei között is lehetséges (például életkori) különbözőség. Szintén nem feltétlenül áll fenn két faj különbözőségének szimmetrikus volta, azaz előfordulhat, hogy dij nem egyenlő dji . Gondolhatunk itt például egyes fajkeresztezési eljárások sémájára, illetve eredményére. Az is előfordulhat, hogy a különbözőség mérőszáma negatív.
Figyelemreméltó körülmény, hogy adott D különbözőségi mátrix esetében Q általában nem egyenletes fajmegoszlás esetén veszi fel maximumát (Izsák - Szeidl, 2002; Champely - Chessel, 2002). Például 14 darufaj DNS-hibridizáción alapuló különbözőségének figyelembe vételével azt kaptuk, hogy a Q diverzitási index értéke az egyenletes megoszlást tükröző (1/14; 1/14; ...; 1/14) relatív gyakoriságegyüttes esetén csak 1,475, míg a (0,405; 0; 0; 0,495; 0; 0; 0; 0; 0; 0; 0,100; 0; 0; 0) relatív gyakoriság-együttes esetén Q értéke 2,465.
Nyilvánvaló, hogy a diverzitási indexek növelését szem előtt tartó természetvédelmi programok céljaira ilyen tulajdonságú index nem jöhet szóba. A genetikai diverzitás kérdéskörében és a populációdinamika egyes területein azonban az eredmény továbbgondolásra érdemes. Hasonló mondható el olyan pszichológiai szituációban, melyben egy kísérleti személy találomra kiválasztott objektumok különbözőségének általa becsült átlagáról számol be.
A szakirodalom hatalmas anyagára tekintettel lehetetlen akár csak közelítően is teljes képet adni az ökológiában folyó diverzitási vizsgálatokról. Fokozza a nehézségeket, hogy lezárt kérdés ebben a témakörben alig van, így a korábbi eredményeket nem lehet átadni a felejtésnek. Másrészt - mint a fentiekből kitűnik - a diverzitási indexek bevezetése több kérdést vetett fel, mint amennyit megoldott. Ez azonban nem szegheti kedvét a statisztikus ökológia e szakterületét művelőknek. Nem mondhatunk le arról, hogy előrehaladjunk a fajközösségek jellemzőinek számszerűsítésében.
Kulcsszavak: diverzitási mérőszámok, fajközösség, kvadratikus entrópia, index-érzékenység
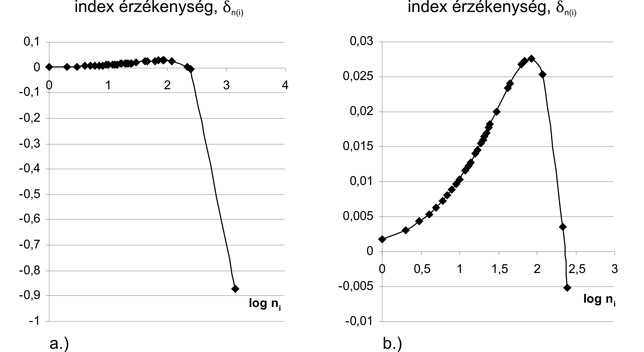
1. Diptera statisztika (Papp, 1992) adatain alapuló Shannon-féle diverzitásra vonatkozó érzékenységi grafikonok.
Irodalom
Champely, Stephane - Chessel, Daniel (2002): Measuring Biological Diversity Using Euclidean Metrics. Environmental and Ecological Statistics. 9, 167-177. http://pbil.univ-lyon1.fr/R/articles/arti106.pdf
Egghe, Leo - Rousseau, Ronald (1991): Transfer Principles and a Classification of Concentration Measures. Journal of the American Society for Information Science. 42, 479-489.
Frigyes Ervin (2002): Struktúra - koncentráció - egyenlőtlenség. Statisztikai Szemle. 78, 598-619.
Izsák János (1996): Sensitivity Profiles of Diversity Indices. Biometrical Journal. 8, 921-930.
Izsák János (2001): Bevezetés a biológiai diverzitás mérésének módszertanába. Scientia, Budapest
Izsák János - Papp László (2002): Sensitivity of Diversity Indices: A Study of Dipterous Assemblages. Community Ecology. 3, 79-86.
Izsák János - Szeidl László (2002): Quadratic Diversity: Its Maximization Can Reduce the Richness of Species. Env. Ecol. Stat. 9, 423-430.
Papp László (2002): Drosophilid Assemblages in Mountain Creek Valleys in Hungary. Diptera: Drosophilidae I. Folia Entomologica 53, 139-153.
Patil, Ganapati P. - Taillie, Charles (1979): An Overview of Diversity. In: Ecologica in Theory and Practice, International Co-Operative Publishing House, Fairland, Maryland, 3-27.
Rao, C. Radhakrishna (1982): Diversity and Dissimilarity Coefficients: A Unified Approach. Theoretical Population Biology. 21, 24-43.
Tóthmérész Béla (1997): Diverzitási rendezések. Scientia, Budapest