
A folyamatot, melynek során gyakorlás vagy tapasztalat következtében tudatosság nélkül megtanulunk érzékelni valamit, amire korábban képtelenek voltunk, perceptuális tanulásnak nevezzük. Mindenki számára jól ismert tény, hogy gyakorlás hatására "finomodnak" az érzékek: a szolfézstanulás finomítja a hallást; a festés és rajzolás a látást; a borkóstolás az ízlelést; a talajtorna a finom koordinációhoz szükséges szomatoszenzoros, testhelyzettel kapcsolatos érzékelést. Alfred Wilhelm Volkmann (1863) már a XIX. században leírta, hogy a tárgyak vizuális felismerése javul gyakorlás hatására - "gyakorlat teszi a mestert". Akkoriban a teljesítményjavulást még kognitívnek, azaz az információfeldolgozás magas szintjein végbemenőnek tulajdonították. Az elmúlt egy-két évtized során elvégzett számos elektrofiziológiai, valamint képalkotó eljárást alkalmazó és pszichofizikai tanulmány azonban kimutatta, hogy a korábbi nézettel ellentétben az elsődleges érzőkérgekben nemcsak a fejlődés korai, ún. kritikus periódusában, de a felnőtt agyban is plasztikus változások zajlanak le. Ennek első indikációja a szomatoszenzoros kéregből származik (Merzenich et al., 1988), de azóta már ismert, hogy az összes szenzoros szubmodalitás úgymint szaglás, ízérzékelés, hallás és látás kérgi területein is megfigyelhető. Retinális léziót követően megváltozik az elsődleges látókéregben lévő receptív mezők helyzete és mérete is (Gilbert et al., 1996, 2000).
A vizuális perceptuális tanulás egy viszonylag új és igen izgalmas területét jelenti a perceptuális tanulásnak. Gondoljunk például a röntgenorvosokra vagy szövettanászokra, akik a vizuális feladat ismételt elvégzése során válnak szakmájuk mestereivé. Pszichofizikai kísérletekben teljesítményjavulás mutatható ki a legtöbb egyszerű vizuális feladatban (vernierdiszkrimináció [McKee - Westheimer, 1978]; textúradiszkrimináció [Karni - Sagi 1991]; sztereoszkopikus mélységlátás [Ramachandran - Braddick 1973]; és vonalorientációk közötti diszkrimináció [Vogels - Orban 1985]). A pszichofizikai kísérletekből az is kiderült, hogy a perceptuális tanulás a vizuális feladatok egy jó részében specifikus a gyakorlásra használt inger alacsony szintű jellegzetességeire. Ez annyit jelent például a textúradiszkriminációs feladatban, hogy egy adott vonaliránnyal (például függőleges) való gyakorlás csak azonos irányú vonalak esetén vezet teljesítménynövekedéshez, a többi vonalirányra egyenként újra kell gyakorolni a feladatot (Fiorentini - Berardi, 1980). Ez az irányspecifikus tanulás arra utal, hogy a tanulás biztosan agykérgi eredetű, mivel az irányszelektivitás először az elsődleges látókéregben jelenik meg. Egy más esetben a látótér adott helyét (például jobb felső negyedét) elfoglaló inger csak az e területen bemutatott ingerlés esetén lesz hatékony a tanulás szempontjából - a látótér más részein újra be kell gyakorolni a feladatot (Karni - Sagi, 1991). Ez a helyspecifikus tanulás azt jelzi, hogy a tanulás valószínűleg az occipitális lebeny alacsony szintű látóterületein belül zajlik, mivel ezekre a területekre jellemző a nagymértékű helyspecificitás (hely alatt a látótér egyes részeit, illetve retinális pozíciókat értve [Karni - Sagi, 1991; Crist et al., 1997]). A vizuális perceptuális tanulás specifikus lehet továbbá például az inger téri frekvenciájára (Fiorentini - Berardi, 1980), a mozgás irányára (Ball - Sekuler, 1987) és a gyakorlásban részt vevő szemre (egy szemmel való tesztelés esetén) (Karni - Sagi 1991; Stickgold et al., 2000b). Ha egy adott feladatban többféle specificitás is jelentkezik, az felhasználható a tanulás neuronális "helyének" meghatározására. Így például ha egy feladat irányspecifikus, akkor biztos, hogy kérgi eredetű a tanulás. Ha ezen túl helyspecifikus is, akkor biztos, hogy alacsony kérgi szinten zajlik, s ha még a gyakorlásban részt vevő szemre is specifikus, akkor biztos, hogy az elsődleges látókéregben megy végbe, ahol a neuronok még részben monokuláris (egy szemmel kapcsolatos) beidegzésűek.
A perceptuális tanulás az idegrendszer plasztikusságát tükrözi. A bekövetkező teljesítményjavulás hatása hosszú távú, hónapokig, akár évekig is eltart (Karni - Sagi, 1993; Julesz - Kovács, 1995). A tanulás azonban nem deklaratív típusú, mivel nem jár tények és események tudatos memorizálásával. Az egyszerű vizuális feladatokban (például textúra diszkrimináció) észlelt perceptuális tanulás időbeli jellemzői hasonlóak a kifinomult motoros feladatok elsajátításához szükségesekhez (Karni et al., 1998): 1) gyors, egyetlen vizsgálati perióduson belüli teljesítményjavulás, 2) a vizsgálati periódusok közötti ún. látens fázis, amely során a teljesítményjavulás lassú, és amely többórás konszolidációt vagy alvást igényel. Uri Polat és Dov Sagi (1995) szerint a gyors komponensben a kéreg magasabb szintjei, míg a lassú komponensben az elsődleges látókéreg vesz részt. Kimutatták, hogy a perceptuális tanulás bekövetkezéséhez időben legalább hat-nyolc óra (Karni - Sagi, 1998), egy rövid délutáni pihenés (Mednick et al., 2001), vagy legalább hatórás éjszakai alvás (Karni et al., 1994; Stickgold et al., 2000a, 2000b) szükséges a gyakorlási szekciók között. Alvásmegvonás a gyakorlás utáni első éjszaka során meggátolja a teljesítményjavulást, még akkor is, ha a következő éjszaka alkalom nyílik az alvásra (Stickgold et al., 2000b). Úgy tűnik tehát, hogy a gyakorlást követő nagyjából harminc óra kritikus az emléknyomok konszolidációját illetően, s hogy ez a konszolidáció nem pusztán az idő múlásával, hanem az alvással magával kapcsolatos. Az egyelőre vitatott azonban, hogy az alvás különböző fázisai, így a gyors szemmozgásokkal járó REM periódus, illetve a mélyalvás fázisa mennyiben, és pontosan hogyan járulnak hozzá a konszolidációhoz (Karni et al., 1994; Stickgold et al., 2000a, 2000b).
Saját kísérleteinkben arra kerestünk választ, hogy egy, a szokásos egyszerű diszkriminációs és vizuális keresési feladatoknál valamivel bonyolultabb feladat gyakorlása során milyen idői paraméterei vannak a tanulásnak. Ebben a feladatban apró építőelemekből kell a látórendszernek összeállítania egy kontúrt (1. ábra).
Az építőelemek ún. Gábor-foltok (Gauss-függvénnyel lesimított, iránnyal rendelkező, sávokból álló szegmentek), melyek az elsődleges látókéreg irányra érzékeny éldetektorait szelektíven aktiválják. A kontúr elemei ugyanolyan Gábor-foltokból álló zajba vannak ágyazva. Ezeknek a foltoknak az irányulása azonban véletlenszerű, azaz nem alkot korrelált mintázatot úgy, mint a kontúr. Az ilyen típusú vizuális ingerek nagyszerűen beváltak az alacsony szintű kérgi vizuális területek integratív kapacitásának vizsgálatában (Kovács, 1996). Az irány információ téri integrációja, az éldetekció és a formalátás közötti feldolgozási lépés, mely lehetővé teszi, hogy a kérgi feldolgozás elején lévő kis receptív mezővel rendelkező idegsejtek lokális jeleiből tárgyakat határoló kontúrok, formák jöjjenek létre. Érdekes módon a feladat, bár igen egyszerűnek tűnik, még öt-hat éves gyerekek számára is nehéz. Méréseink szerint az emberi perceptuális fejlődés során legalább tíz évet vesz igénybe, hogy ez az ún. kontúrintegráció felnőtt szinten működjön (Kovács et al., 1999). A "kritikus periódus" tehát ebben az esetben igen hosszú. Eddigi eredményeink azt mutatják, hogy a feladatban részt vevő agyi mechanizmusok nagy valószínűséggel az elsődleges látókéregben helyezkednek el (Kovács 1996; Giersch et al., 2000). Gyerekek és felnőttek egyaránt fejlődést mutatnak, ha három egymást követő napon gyakorolhatják a feladatot, és ez a fejlődés specifikus az ingert meghatározó jellemzőre (irány, illetve szín által meghatározott kontúrok esetén a tanulás nem transzferál a két jellemző között [Kovács et al., 1999]). A specifikusság további indikáció az alacsony szintű kérgi feldolgozásra. Jelen tanulmányunkban új eredményeinket közöljük, melyek demonstrálják, hogy a kontúrintegrációs feladatban mutatkozó tanulás idői jellegzetességei hasonlóak az egyszerű vizuális feladatok során mutatkozó tanulás idői jellegzetességeihez, s hogy itt is "aludni kell egyet" a gyakorlásra.
Huszonnégy normál felnőtt naiv alanyt vizsgáltunk. Minden alany öt egyforma hosszú gyakorlási perióduson esett át, de a gyakorlás időbeli eloszlása különbözött, s eszerint osztottuk három csoportba őket: 1. csoport: az öt gyakorlási periódus 3,5 óra alatt zajlott le, 2. csoport: az öt gyakorlási periódus tíz óra alatt zajlott le, 3. csoport: az öt gyakorlási periódus öt nap alatt zajlott le. Az egyes csoportok jellemzőit lásd az 1. táblázatban. Az alanyok közül senki nem szenvedett amblyopiában vagy kancsalságban, a látásélesség korrekcióval vagy anélkül teljes volt. A látásélesség vizsgálata Snellen-táblával történt.
A vizsgálatban használt ábrák generálása a korábban leírtakhoz hasonlóan történt (Kovács et al., 1999). A Gábor-foltok téri frekvenciája 5 c/deg, míg kontrasztja 95 % volt. Ebben a közleményben a kontúrintegrációs feladatnak egy új verzióját alkalmaztuk. Az ábrákon kolineáris Gábor-foltokból álló kontúr és ugyanezen foltok random orientációjú halmazából álló háttér található (1. ábra). Az ábrák mindegyikén a kontúr egy elfektetett tojást rajzol ki. Az alanyok feladata az volt, hogy az egér megfelelő gombjainak lenyomásával jelezzék, hogy a tojás hegyesebbik vége balra vagy jobbra mutat-e. Az ábrákat számítógép képernyőjén jelenítettük meg, amelyet az alanyok 40 cm távolságból, ülve néztek, egy mesterséges fénnyel megvilágított szobában. Az ábrák a vizsgálati távolságból függőlegesen 19,93°, míg vízszintesen 26,57° szög alatt voltak láthatóak. A monitor átlagos luminenciája 21,5 cd/m2 volt. Az ábrák két másodpercig kerültek kivetítésre. Ez alatt az idő alatt kellett az alanyoknak a választ megadni. Az ábrák közé egy másodperc hosszúságú szünetet iktattunk be, amely során a képernyő egyszínű szürkére változott, és annak közepén egy fixációs kereszt jelent meg. Az ábrákon a háttérelemek és a kontúrelemek távolságának hányadosa, az általunk korábban D-nak nevezett érték, normál felnőtt kontúrintegrációs küszöbszintet jóval meghaladó, állandó volt (D=0,9). A D értékének állandó szinten tartása lehetővé tette a kérgi neuronok közötti interakciók kizárólagos vizsgálatát, és nem adott lehetőséget az ábrán sűrűségváltozásra a Gábor-foltok között. Az ábrákon kizárólag a kontúrelemek orientációja változott kollineáristól (0°) 28°-ig, hét különböző nehézségi szinten: 0°, 7-8°, 11-12°, 15-16°, 19-20°, 23-24°, 27-28° (2. ábra). Egy-egy nehézségi szinten belül a kontúrelemek orientációját random módon változtattuk 0,1 fokonként, a két feltüntetett érték között. Az alanyok negyven ábrát néztek meg 4x10-es csoportokban az egyes nehézségi szinteken. Ezen ábrák egyik felén a tojás hegye jobbra, a másik felén balra mutatott, de a jobb, illetve bal oldalra mutató ábrák bemutatása random sorrendben történt. Az egyes szintek fokozatosan nehezedő sorrendben követték egymást. A tesztelést binokulárisan végeztük.
Egy gyakorlási periódus hossza körülbelül 20-25 perc volt. Küszöbteljesítménynek a 75 %-os helyes választ tekintettük. Az alanyok a tesztelés megkezdése előtt egy tíz ábrából álló "bemelegítő" sorozatot néztek meg a legkönnyebb nehézségi szinten. A tesztelés akkor kezdődött, amikor az alanyok megbízható módon legalább 80 %-os teljesítményt nyújtottak ezzel a "bemelegítő" sorozattal. A tesztelés során nemcsak a helyes válaszok, de a válaszadás elmaradása is regisztrálásra került. A vizsgálatban résztvevők között nem volt szükség a válaszadás elmaradása következtében a végleges teljesítmény korrekciójára. Egyetlen alanynál sem haladta meg ugyanis annak értéke a 2,5-5 %-ot a vizsgálat teljes időtartama során. Az egyes nehézségi szinteken a gyakorlási periódus során elért átlagos teljesítményt pszichometriai görbén ábrázoltuk. Az alanyoknál a küszöbértéket logisztikai függvénynek az adatpontokra való fektetésével kaptuk meg. A végleges küszöbérték-adatokat statiszikailag az egyes csoportokon belül egyszempontos ANOVA (ismételt mérések), valamint páronkénti összehasonlítás (LSD korrekcióval) módszerével elemeztük. A legelső periódust az egyes csoportok között egyszempontos ANOVA (Tukey HSD Post-hoc teszt) segítségével hasonlítottuk össze.
A csoportok között a legelső gyakorlási periódusban nyújtott teljesítményben szignifikáns különbség nem mutatkozott (p>0,05 - egyszempontos ANOVA). Az első csoportban (három és fél óra gyakorlás) és a második csoportban (tíz óra gyakorlás) az egyes gyakorlási szekciók között statisztikailag nem volt eltérés (p>0,05 - ANOVA ismételt mérések) (3. ábra). Az ötnapos, harmadik csoportban szignifikáns napi hatást találtunk. Szignifikáns eltérés mutatkozott az első és a negyedik, valamint az első és az ötödik nap között (F=4,385; df=1,936; p=0,035 - ANOVA , LSD). A napszakok nem voltak hatással a teljesítményre az 1. és a 3. csoportban.
Bár a három csoport ugyanannyi időt töltött gyakorlással (ötször fél órát), az eredmények azt mutatják, hogy csak abban a csoportban jött létre szignifikáns tanulás, amelyik a gyakorlási periódusok között hozzájutott legalább egy éjszaki alváshoz. Önmagában a gyakorlási periódusok között eltelt idő nem hozott változást, tehát ez a fajta konszolidáció, legalábbis a mért időtartományban, nem egyszerűen az idő, hanem az alvás függvénye. Bár az is lehetséges, hogy az alvás és a gyakorlási periódusok időbeli megoszlása együttesen játszanak szerepet. Eredményeink összhangban állnak más, egyszerűbb vizuális feladatok során mutatkozó tanulási eredményekkel (például Karni - Sagi, 1993; Mednick et al., 2001; Karni et al., 1996; Stickgold et al., 2000a, 2000b).
Összevetve saját korábbi eredményeinkkel, melyek a tanulás ingerspecifikusságára mutattak rá ebben a feladatban (Kovács et al., 1999), arra a következtetésre jutunk, hogy a vizuális téri integrációban részt vevő mechanizmusok felnőttkorban is plasztikusak, s hogy ez a plaszticitás az alacsony szintű kérgi vizuális területek sajátja.
Kulcsszavak: perceptuális tanulás, konszolidáció, alvás, kontúrintegráció
1. ábra * A perceptuális tanulási feladatban alkalmazott kontúrintegrációs inger. A kontúr kolineárisan irányuló Gábor-foltokból áll, melyek között a távolság valamivel kisebb, mint a háttérelemek közötti átlagos távolság. Így az egyetlen jelzés, ami lehetővé teszi a kontúr detekcióját, a kontúrt alkotó Gábor-foltok egymáshoz viszonyított iránya. Ennek kódolását az irányérzékeny agykérgi idegsejtek közötti irányfüggő anatómiai összeköttetések végzik (valószínűleg az elsődleges látókéregben). Így ezzel az ingerrel az irányfüggő összeköttetések szelektíven erősíthetők a gyakorlás során.
Csoportok Átlagéletkor (év) Teljes tesztidő Férfi:nő arány 1. (n=8) 24,5 3,5 óra 4:4 2. (n=8) 29,75 10 óra 3:5 3. (n=8) 28,75 5 nap 4:4
1. táblázat * A csoportok általános jellemzői. A táblázat csoportonként mutatja a résztvevők számát, átlagos életkorát - években, a gyakorlás teljes időtartamát, valamint a férfiak és nők megoszlását.

2. ábra * A perceptuális tanulási feladatban alkalmazott kontúrintegrációs inger egyre nehezedő változatai. A feladatban a képeken lévő, fekvő tojás alakú kontúrokról el kellett dönteni, hogy a tojás hegyesebbik vége jobbra vagy balra mutat-e. A feladat nehézségét a kontúrelemeknek a kontúr képzeletbeli vonalához képest való elforgatásával változtattuk, és mértük, hogy mi az a legnagyobb elforgatás, ami mellett még elvégezhető a feladat (küszöb). Az elforgatás mértéke a mintaként bemutatott ábrák alatt látható. Kísérleti alanyaink átlagos küszöbe 11-12° körül volt (középső ábra). 23-24 fokos elforgatás már lehetetlenné teszi a feladat megbízható módon való végrehajtását (alsó ábra). Legjobban tanuló csoportunk átlagosan 2° javulást ért el öt nap alatt (ötször fél óra abszolút gyakorlási idő mellett).
1. periódus 2. periódus 3. periódus 4. periódus 5. periódus 1. csoport 12,99 13,55 14,06 13,75 13,23 2. csoport 10,9 11,02 11,13 10,43 11,76 3. csoport 12,57 13,36 13,16 14,01 14,58
2. táblázat * Átlagos küszöbértékek. A táblázatban a gyakorlási periódusokban kapott, az egyes csoportokra jellemző, átlagos küszöbértékeket tüntetjük fel fokokban.
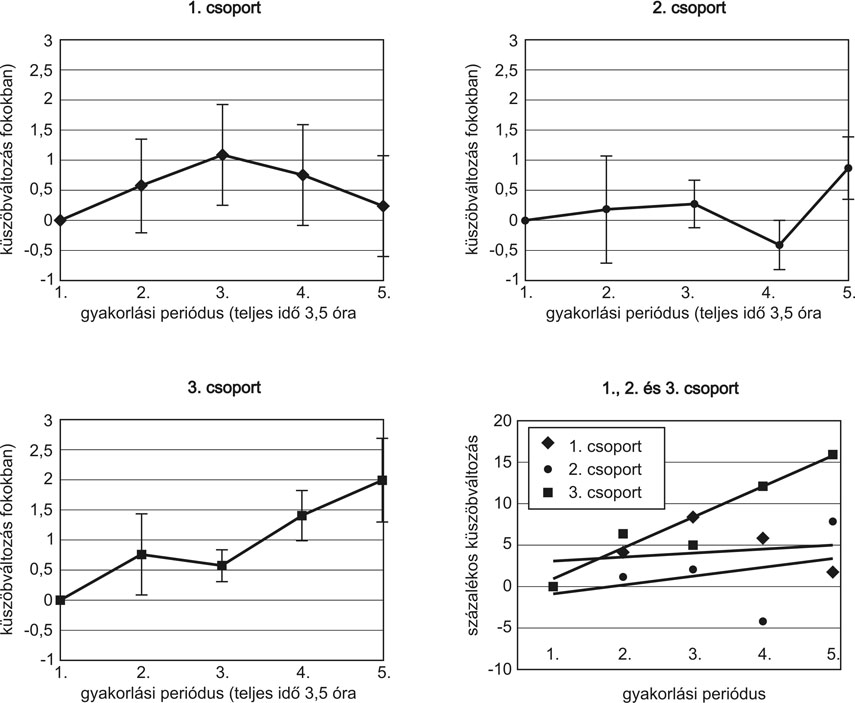
3. ábra * A három csoport egyéni (a, b, c) és összevont tanulási görbéje (d). Az egyéni tanulási görbék fokokban mért változást mutatnak a gyakorlási periódusok előrehaladtával. Mindhárom csoport öt gyakorlási perióduson ment keresztül, de a periódusok idői eloszlása eltért csoportonként. Az első csoport három és fél óra, a második tíz óra leforgása alatt végezte el az öt gyakorlási periódust. A gyakorlási idő megnyújtása önmagában nem hozott szignifikáns tanulási hatást. A harmadik csoport gyakorlási periódusai között egy-egy nap telt el. Ők a negyedik, ötödik napra már szignifikánsan jobb teljesítményt értek el, mint az első napon.
A három csoport teljesítményét összehasonlító ábrán (d) százalékban fejeztük ki a küszöbváltozást, és a csoportok görbéit lineáris regressziós egyenessel helyettesítettük, hogy a tanulás tendenciája jobban láthatóvá váljon. Ebben az ábrázolásban világos, hogy csak a harmadik csoport teljesítménye mutat konzisztens javulást.
IRODALOM
Ball, Karlene - Sekuler, Robert (1987). Direction-specific Improvement in Motion Discrimination. Vision Research. 27, 953-965
Crist, Roy E. - Kapadia, M. K. - Westheimer, G. - Gilbert, C. D. (1997). Perceptual Learning of Spatial Localization: Specificity for Orientation, Position and Context. Journal of Neurophysiology. 78, 2889-94
Fiorentini, Adriana - Berardi, Nicoletta (1980). Perceptual Learning Specific for Orientation and Spatial Frequency. Nature. 287, 43-44
Giersch, Anne - Humphreys, G. - Boucart, M. - Kovács I. (2000). The Computation of Occluded Contours in Visual Agnosia: Evidence for Early Computation Prior to Shape Binding and Figure-Ground Coding. Cognitive Neuropsychology. 17, 731-759
Gilbert, Charles D. - Das, A. - Ito, M. - Kapadia, M. - Westheimer, G. (1996). Spatial Integration and Cortical Dynamics. Proceedings of the National Academy of Sciences of the USA. 93, 615-622
Gilbert, Charles D. - Ito, M. - Kapadia, M. - Westheimer, G. (2000). Interactions between Attention, Context and Learning in Primary Visual Cortex. Vision Research. 40, 1217-1226
Julesz Béla - Kovács Ilona (1995). Introduction. In: Maturational Windows and Adult Maturational Windows and Adult Cortical Plasticity. SFI Studies in the Sciences of Complexity. Vol. XXIII. Addison-Wesley, Reading, MA, Xxix-Xxiv.
Karni, Avi - Sagi, Dov (1991). Where Practice Makes Perfect in Texture Discrimination: Evidence for Primary Visual Cortex Plasticity. Proceedings of the National Academy of Sciences of the USA. 88, 4966-4970
Karni, Avi - Sagi, Dov (1993). The Time-Course of Learning a Visual Skill. Nature. 365, 250-252
Karni, Avi - Tanne, D. - Rubenstein, B. S. - Askenasy, J. J. - Sagi, D. (1994). Dependence on REM Sleep of Overnight Improvement of a Perceptual Skill. Science. 265, 679-682
Karni, Avi - Meyer, G. - Rey-Hipolito, C. - Jezzard, P. - Adams, M. M. - Turner, R. - Ungerleider, L. G. (1998). The Acquisition of Skilled Motor Performance: Fast and Slow Experience-Driven Changes in Primary Motor Cortex. Proceedings of the National Academy of Sciences of the USA. 95, 861-68
Kovács Ilona (1996). Gestalten of Today: Early Processing of Visual Contours and Surfaces (A Review). Behavioural Brain Research. 82,1-11
Kovács Ilona - Kozma P. - Fehér Á. - Benedek, Gy. (1999). Late Maturation of Visual Spatial Integration. Proceedings of the National Academy of Sciences of the USA. 96, 12204-12209
McKee, Suzanne P. - Westheimer, Gerald (1978). Improvement in Vernier Acuity with Practice. Perception & Psychophysics. 24, 258-262
Mednick, Sara C. - Atienza, M. - Cantero, J. L. - Nakayama, K. - Stickgold, R. (2001). Benefit of Nap Is Retinally Specific. VSS (Vision Sciences Society) Abstract
Merzenich, Michael M. - Recanzone, G. - Jenkins, W. M. - Allard, T. T. - Nudo, R.J. (1988). Cortical Representational Plasticity. In: Rakic, P. - Singer, W. (eds.) Neurobiology Of Neocortex. Wiley, New York, 41-67
Polat, Uri - Sagi, Dov (1995). Plasticity of Spatial Interactions in Early Vision. In: Julesz Béla - Kovács Ilona (eds.) Maturational Windows and Adult Cortical Plasticity. SFI Studies In The Sciences of Complexit. Vol. 24. Addison-Wesley, Reading, MA.
Ramachandran, Vilayanur S. - Braddick, Oliver (1973). Orientation-Specific Learning in Stereopsis. Perception. 2, 371-376
Stickgold, Robert - James, LaTanya - Hobson, J. Allan (2000a). Visual Discrimination Learning Requires Sleep after Training. Nature. 3, 1237-1238
Stickgold, Robert - Whidbee, D. - Schirmer, B. - Patel, V. - Hobson, J. A. (2000b). Visual Discrimination Task Improvement: A Multistep Process Occurring during Sleep. Journal of Cognitive Neuroscience. 12, 246-254
Vogels, Rufin - Orban, Guy A. (1985). The Effect of Practice on the Oblique Effect in Line Orientation Judgements. Vision Research. 25, 1679-1687
Volkmann, Alfred Wilhelm (1863). Physiologische Untersuchungen Im Gebiete Der Optik. Breitkopf Und Härte, Leipzig