Szociális kogníció
Az etológusok már a kezdetektől fogva nagy érdeklődéssel vizsgálták a csoportban élő állatok szociális viselkedését. A récefélék udvarlási szokásai, a tüskés pikók agresszív magatartásformái vagy a sirályok egyedi felismerése mind részei az egyes fajok szociális magatartási formáinak. A hetvenes években a szociobiológia megjelenésével újabb lendületet kapott a szociális magatartásformák tanulmányozásának kutatása, amikor előtérbe került, hogy az egyes fajok többnyire genetikailag meghatározott szociális magatartásformái milyen mértékben járulnak hozzá a faj túléléséhez. A szociális magatartásformák iránti érdeklődés harmadik szakasza talán a nyolcvanas évekre tehető, amikor a kognitív pszichológiai "forradalom" hatása túllépett a tanulási folyamatok vizsgálatán, és a kognitív megközelítést a viselkedésszerveződés egészére általánosították. Ekkor születik meg a kognitív etológia mint önálló tudományterület, illetve kezdődnek azok az összehasonlító kutatások, amelyek csimpánzok és az ember viselkedését összevetve igyekeznek evolúciós perspektívában vizsgálni a szociális viselkedés egyes komponenseit. Új lendületet kapnak a csecsemő- és gyermekvizsgálatok is, hiszen a szociális magatartásformák egyedfejlődésének megértése alapvető az epigenetikus folyamatok (a genetikai információ és a környezet közötti kölcsönhatás) értelmezéséhez.
Az idők során a szociális kogníció fogalma is némi változáson ment át. Kezdetben elsősorban az elmeteóriához (lásd később) közvetlenül kapcsolódó jelenségeket sorolták ide, ma azonban a legtöbb szerző sokkal szélesebb értelemben használja ezt a kifejezést, felismerve, hogy az állatok és az ember környezetében előforduló "objektumok" alapvetően kétfélék lehetnek: passzívak és aktívak. A passzív objektumok maguktól nem mozognak, és nem reagálnak az állat viselkedésére, leszámítva a külső behatások vagy a fizikai törvényszerűségek miatt bekövetkező állapotváltozásokat, például a fán lógó alma leesik a földre. Az aktív objektumok rendelkeznek az önálló mozgás képességével, illetve képesek komplex interakcióba lépni az egyeddel. Az effajta belső autonómia az élőlényekre jellemző, melyek az egyed szempontjából két fő csoportba sorolhatók: fajtársak, illetve nem-fajtársak (ezek közül is külön jelentőséggel bírnak a fajra veszélyt jelentő ragadozók, illetve a potenciális prédafajok). Jelenleg széles körben elfogadott meghatározás szerint a szociális kogníció alatt mindazokat a viselkedésbeli képességeket értjük, amelyeket az egyed a fajtársakkal való együttélés során használ. Így a szociális kogníció fogalmába tartozik a fajtárs felismerése és a fajtársak közötti kötődés éppúgy, mint a fajtársakkal való kommunikáció vagy kooperáció. A fentieket úgy is összefoglalhatjuk, hogy a szociális kogníció azokat a kognitív folyamatokat és azok magatartásbeli megnyilvánulásait foglalja magában, amelyek a szociális környezetre vonatkoznak, szemben a többi, a fizikai környezetre vonatkozó magatartásformákkal. Ebben az értelemben is logikus a szociális kogníció széles körben vett értelmezése, hiszen e megközelítés szerint az első lépes az adott objektum beazonosítása ("passzív" vagy "aktív"), ehhez pedig az állati elmének szüksége van speciális felismerő rendszerre, amely több lényeges vonásban eltér a "passzív" tárgyakra vonatkozó felismerő rendszertől.
A felismerés különbségein túl talán a legfontosabb eltérés, hogy az "aktív" ágensek esetében az egyednek rendelkeznie kell olyan képességekkel, melyek segítségével képes bejósolni az objektum viselkedését, illetve ennek ismeretében képes felkészülni bizonyos jövőbeli akciókkal való szembesülésre (anticipáció). Egy ragadozóval való találkozáskor vagy egy fajtárssal való interakcióban az az egyed lesz sikeresebb, amely jobban alkalmazza a jóslás, illetve az anticipáció módszerét.
Érdemes azonban már az elején megemlíteni, hogy az "aktív" objektumok (ágensek) rendelkeznek az egyed szempontjából még egy tulajdonsággal, amely hozzájárul a velük való interakció komplex mivoltához. Eddig ugyanis az aktivitást csak az egyedhez viszonyítva tárgyaltuk (diadikus interakció), pedig ezek az ágensek természetükből adódóan egymással is folyamatos interakcióban lehetnek oly módon, hogy ennek a szóban forgó egyed csak megfigyelő részese (triadikus interakció). Ebből következően az egyed oly módon is juthat az "aktív" ágensekre vonatkozó információ birtokába, hogy ő maga nem közvetlenül vesz részt az interakcióban. Rangsorviták esetén például előnyös tudni, hogy ki milyen támogatókra számíthat. Egyes majomfajoknál például az egymással baráti viszonyban levő egyedek többet kurkásszák egymást. Ily módon a csapattagok kurkászási interakcióiból kikövetkeztethetők azok a személyes preferenciák, amelyeket figyelembe lehet venni egy csapattárs elleni fellépés esetén.
Elmeteória: vajon gondolkodik-e egy csimpánz mirólunk?
A hetvenes években David Premack és Guy Woodruff (1978) egy első hallásra igencsak furcsa kérdésre keresték a választ: vajon mit gondolhat egy ember által nevelt csimpánz az emberről? A kérdés megválaszolására több kísérletet is terveztek. Az egyik ilyen kísérletsorozatban Sarah, egy nősténycsimpánz egy videofelvételen azt láthatta, hogy egy szobában egy ember szeretne valamilyen problémát megoldani, például felugrálva szeretne hozzájutni egy fellógatott banánhoz, de nem éri el. A film megtekintése után Sarahnak három különböző tárgy képét (például létra, labda, ablak) mutatták, amelyből egyet kellett kiválasztania. Érdekes módon a csimpánz meglehetős biztonsággal választotta ki azt a tárgyat, amelyik valóban szükséges volt a kérdéses probléma megoldásához. Az eredmények alapján Premack és Woodruff úgy érveltek, hogy Sarah képes felismerni a másik akciójának a célját, azaz képes szándékot tulajdonítani az embernek ("a másik akar valamit"). Más szóval képes felismerni a másik egyed mentális állapotát, és a saját viselkedését ennek megfelelően változtatni. Eszerint elképzelhető, hogy a csimpánz elméje rendelkezik olyan speciális képességgel, amely a másik elmeállapotát felismeri, illetve reprezentálja. Részben alternatív lehetőség, hogy a csimpánz nem a másik szándékát ismeri fel, hanem korábbi tapasztalatai alapján oksági alapon következtetett az adott helyzetre vonatkozóan ("ha fellógatott banánt látsz, akkor létra kell a megszerzéshez").
A történet érdekessége, hogy a másik elméjének reprezentációs problémája épp a csimpánz-ember viszonylatában merült fel először, amikor akkortájt még az sem volt egészen világos, hogy például az ember egyedfejlődésének mely szakaszában jut az elmeteóriához szükséges képességek birtokába. Sajnos az azóta eltelt évek során a csimpánzok elmeteóriával kapcsolatos képességeire elsősorban módszertani problémák miatt nem sikerült egyértelmű bizonyítékot találni, míg az eltelt idő alatt viszonylag alapos ismereteket sikerült szerezni a gyerekek elmeteóriára érintő képességeiről.
Asszociáció, reprezentáció, modularitás
Az elmeteóriát érintő kísérletek kezdetén kibontakozott (és változó hevességgel ma is folyik) a tanulási folyamatok esetében már korábban lezajlott vita, vajon ezek a szociális képességek mennyiben tekinthetők pusztán egy komplex tanulási képesség megnyilvánulásának, azaz kellő tapasztalat (tanulás) nyomán az egyed lényegében egyszerű asszociációs mechanizmusok révén sajátítja el, hogy miképp kell egy adott helyzetben viselkedni. A probléma jobb megértése kedvéért nézzünk röviden egy másik idevágó kísérletet: Daniel J. Povinelli és munkatársai (1990) egy kooperáción alapuló kísérleti helyzetben arra tanítottak meg csimpánzokat, hogy két edény közül válasszák ki azt, amelyikbe a kísérletvezető előzőleg egy darab táplálékot rejtett. A kísérlet következő szakaszában az alanyok a rejtést nem láthatták, de a táplálék helyét a kísérletvezető mutatással jelezte. Mikor a csimpánzok már ebben a helyzetben is jól választottak, a kísérletvezető rejtését egy másik ember is láthatta. Innen kezdve azonban nem a kísérletvezető jelzett, hanem a "látó-tudó" ember, illetve egy másik ember, aki a rejtés idején nem volt a helyiségben, így nem tudhatta, hol van a táplálék. A két ember egyszerre mutatott valamelyik edényre (a "tudó" mindig a helyesre, a "tudatlan" mindig a helytelenre), és a csimpánznak ez alapján kellett választania. Többpróbás sorozatban vizsgálva, úgy tűnt, hogy a csimpánzok felismerték a helyzetet, és választásukat a rejtéskor jelen lévő ember jelzése alapján tették meg. Ezt követően a kísérletet úgy módosították, hogy mindkét ember a helyiségben maradt, de a rejtés előtt a "tudatlan" ember egy vödröt tett a fejére. Povinelliék úgy érveltek, hogy amennyiben a csimpánzok ebben az esetben helyesen választanak, akkor képesek a másik elmeállapotának figyelembe vételére. Több próba eredményét figyelembe véve úgy tűnt, hogy a négy egyedből három valóban az elvárásnak megfelelően választott, azaz viselkedésük alátámasztotta az elmeteóriából következő jóslásokat. Sokan azonban felvetették, hogy a sok tréning, illetve az előzetes tapasztalat más csimpánzokkal illetve emberekkel önmagában is elegendő ilyen képesség kialakulásához, s a jelenség könnyen magyarázható asszociációs tanulással, hiszen a csimpánz megtanulhatta, hogy például az az ember jelez helyesen, akinek az arca látható a rejtés során. Valóban, az új típusú próbák kezdetén a csimpánzok véletlenszerűen választottak, s a helyes válaszok aránya csak fokozatosan emelkedett a próbák során. Érdemes megjegyezni, hogy azóta kiderült: a csimpánzok sokkal jobb teljesítményt nyújtanak hasonló felépítésű, de kompetíción alapuló helyzetekben, amelyek jobban megfelelnek a csimpánzok természetben is megfigyelhető szociális rendszerének. A fentiekben leírt kooperációs készségre épülő helyzetben a kutyák jobb teljesítményre képesek (például Soproni et al., 2001), ami talán kevésbé meglepő, ismerve a kutya-ember (gazda) közötti kapcsolat kooperatív mivoltát.
Bár a vita máig sincs teljesen lezárva, a szociális kogníció kutatóinak többsége mégis inkább hajlik arra, hogy a szociális kogníció nem egyszerűen a fejlettebb asszociációs képességen alapszik, hanem feltételezik, hogy a háttérben az evolúció során erre a feladatra specializálódott, részben genetikailag is meghatározott agyi struktúrák állnak, amelyek működésének következménye az állati (és emberi) agyban a szociális térről kialakított, reprezentációkon alapuló környezeti modell. Az elképzelés lényege, hogy az agyban léteznek olyan funkcionális működési egységek ("modulok"), amelyek feladata a szociális kogníció egyes jól körülírt elemeinek reprezentálása. A moduláris felépítés feltételezi, hogy az egyes funkciók szelektíven meghibásodhatnak (lásd például autizmus), a modul működése gyors és automatikus, nem kell hozzá tudatos agyműködés, illetve irányított figyelem, a faj összes egyedére jellemző (genetikailag meghatározott képesség) és az egyedfejlődés adott szakaszában jelenik meg. A modularitásra vonatkozó hipotézisből nem következik szükségszerűen, hogy az egyes modulok megfeleltethetők egy adott körülhatárolt idegi struktúrának, ennek ellenére (jobb híján) számos kutató ennek igazolására törekszik. A teljesség kedvéért meg kell jegyezni, hogy a modularitásra vonatkozó elképzelés nem zárja ki asszociatív mechanizmusok létezését, azonban a genetikailag meghatározott modulok feltételezése azt jelenti, hogy minden faj esetében az egyed egy már előre részben "huzalozott" reprezentációs rendszerrel rendelkezik, mely bizonyos értelemben adott kategóriák szerint bontja fel a szociális környezetet, ami meghatározza a későbbi értelmezési keretet. Például egyre több adat utal arra, hogy a másik tekintetirányának felismerésében döntő szerepet játszik a szemek nézési iránya. A temporális lebenyben a kutatók leírtak olyan neuronokat, amelyek különösen érzékenyek a másik szemeinek irányulására. Ezek alapján feltételezik, hogy a tekintetfelismerést egy külön erre a feladatra specializált neuronegyüttes ("modul") végzi. Végső soron megállapítható, hogy általános értelemben a szociális kogníció olyan genetikailag meghatározott struktúrákon alapul, amelyek folyamatos interakcióban fejlődnek ki a környezettel és kultúrával.
A szociális kogníció általános modellje
Az utóbbi években számos szerző igyekezett egy egységes modellt szerkeszteni, amely általános formában mutatja be e képesség főbb komponenseit. Míg a kutatók egy része elsősorban viselkedési adatokon alapuló modellek kifejlesztésén fáradozott, addig egy másik csoport igyekezett a modell egyes komponenseinek meghatározását egyben idegi struktúrákhoz kötni. A teljes értelmezéstől még messze vagyunk, de egyre inkább felmerül az igény az eltérő megközelítésen alapuló, de sok tekintetben hasonló modellek egységesítésére. Az alább felvázolásra kerülő modell több szerző elképzeléseit igyekszik ötvözni. A modellnek három fő összetevője van: (1) az ágens viselkedésének felismerése és dekódolása különböző szinteken, (2) az ennek következtében létrejövő speciális reprezentációk, illetve azok a folyamatok, amelyek (3) egyedfejlődési értelemben kapcsolatot teremtenek, és lehetővé teszik, hogy a felismerés következményeképpen speciális reprezentációk jelenjenek meg.
Viselkedési és idegrendszeri adatok egyaránt alátámasztják, hogy egy ágens felismerése több szinten zajlik. Neurális szinten akkor beszélhetünk felismerésről, ha sikerül találni olyan neuronokat, amelyek speciálisan érzékenyek bizonyos külső ingerek megjelenésére. Az ilyen "felismerő" neuronok kutatói hangsúlyozzák, hogy itt sokszor nem "igen-nem" válaszokról van szó (bár erre is akad példa), hanem relatív specializációról. Vizuális modalitásban az ilyen típusú kísérletek úgy zajlanak, hogy egysejtes elvezetés mellett a kísérleti egyednek (például makákók) különféle képeket mutatnak, amelyek egy sajátságban invariánsak, például mindegyiken egy szemöldökét ráncoló (agresszív jelzés) fajtárs látható. Megfelelő kontrollképek segítségével kimutatható, hogy a makákók amigdalájában léteznek olyan sejtek, amelyek speciálisan érzékenyek az ilyen arcokra. Kísérleti eredményekre támaszkodva egyes kutatók feltételezik (Brothers - King, 1992), hogy ez a felismerés több szinten valósul meg. A legalacsonyabb szintet a testrészek mozgásának felismerése jelenti. Így például a makákók mediális amigdalájában találtak olyan neuront, amely a lépés során látható lengő combmozgásra volt érzékeny, és ráadásul még preferenciát is mutatott az előre történő mozgásra. Kontrollkísérletekben kiderült, hogy ez a neuron nem tüzel, ha a videón bemutatott filmrészleten egy olyan előremozgó egyed látható, amelynek lábait nem mutatja a felvétel. A legmagasabb felismerési szintre példa a "megközelítésnek" mint akciónak felismerésére specializált neuron, amely akkor tüzelt hevesen, ha egy makákó megközelített egy másik egyedet. A jelenség úgy is értelmezhető, hogy az idegsejt nem pusztán a mozgást, hanem annak célját is "reprezentálja", ami feltehetően a szándékosság magasabb szintű értelmezésének első lépése. Szintén ide sorolhatók azok az idegsejtek, amelyek komplex arcmimikát ismernek fel. Sikerült azonosítani olyan neuronokat az amigdalában, amelyek a komolyabb agressziót jelző szemöldökráncolásra vagy az agresszió enyhébb formáját előrevetítő (alternáló rá-, majd elnézés) jelzésre mutattak szeletív aktivitást. A kontrollként használt felvételek, melyeken egy táplálékot rágó vagy fejét forgató majom volt látható, nem bizonyultak hatásosnak.
A felismerő neuronok sok szempontból speciális csoportját képezik azok, amelyek a szemek, illetve a fej mozgását "figyelik" (1. ábra). Az elmeteória szempontjából az ilyen viselkedési formáknak azért van döntő jelentőségük, mert ezek képezik az alapját annak, hogy a másik viselkedésének megfigyeléséből annak mentális állapotára következtethessünk. A másik tekintetének felismerése még egy járulékos érdekességgel bír: a másik tekintetének iránya erős késztetést ébreszt a megfigyelőben a tekintet irányának követésére. Ez a reflexszerű tekintetkövetés már tizennégy-tizennyolc hónapos kortól megjelenik (egyesek szerint még korábban), és felnőttkorban is megmarad. Sőt, a kísérletek tanúsága szerint gátolni igen nehéz. Kísérleti személyek még akkor is követik a tekintetet, ha előzőleg felszólítják őket arra, hogy ne figyeljenek rá (Langton et al., 2000).
A tekintetfelismerő neuronok fő lokalizációja a sulcus temporalis superior (elülső-oldalsó agykérgi terület; ún. STS régió), amely azonban gazdag kölcsönös kapcsolatban áll az amigdalával. További kísérletek alapján kiderült, hogy mások tekintetirányának meghatározásában az agy két fő jelzésre támaszkodik: (1) a pupilla helyzete a szemfehérje látható részéhez képest, illetve a (2) fej irányultsága (amelynek dekódolásában az orrirány játszhat fontos járulékos szerepet). Míg egyes szerzők a szemirányultság domináns voltát hangsúlyozzák (Perrett et al., 1985), addig újabb adatok a szem és a fej helyzetének párhuzamos kiértékelését tartják valószínűnek, és szerintük a viselkedés értelmezése a két jelzőrendszer állapotainak eredőin alapul (Langton et al., 2000). Az ágens viselkedésének dekódolása adja az elme számára a szándék felismeréséhez, illetve reprezentációjához szükséges információt, amelyre szükség van ahhoz, hogy a megfigyelő az ágens viselkedését, valamint annak a környezeti irányultságát egy célra törekvésként értelmező mentális modellbe helyezze (2. ábra, Baron-Cohen, 1994).
Például ennek a rendszernek egyik speciális működési formája, amikor láthatóan élettelen tárgyak, például monitoron egymással mozgási kapcsolatba kerülő geometriai figurák viselkedését (háromszög, kör stb.) élőlényekre alkalmazott verbális terminusokban írjuk le: A nagy háromszög üldözi a pici kört, ami menekül előle. Ahelyett, hogy azt mondanánk: a nagy háromszög követi a pici kört.
A modell másik fontos része a másik elmeállapotának reprezentációjára vonatkozik. Ha elmeállapot-reprezentációról beszélünk, akkor mindenki általában a másik elméjében meglévő környezetre vonatkozó információ reprezentációjára gondol, pedig a kérdésnek van egy másik fontos aspektusa is. Vegyük észre, hogy általános értelemben az elme nemcsak a környezetre vonatkozó információt tárolja, hanem saját maga is egy adott állapotban van. Egy külső megfigyelő szempontjából tehát az ágens reprezentációjában nemcsak annak környezetre vonatkozó állapotát kellene reprezentálnia, hanem a másik belső állapotát is. Leslie Brothers és Brian King (1992) eszerint megkülönbözteti az elmeállapot reprezentáció "hideg" és "meleg" aspektusát. Míg ez előbbi mechanizmus arra szolgál, hogy az ágens múltbéli illetve jelen viselkedése alapján annak elméjének környezetre vonatkozó információját reprezentálja, addig az utóbbi mechanizmus feladata az ágens belső motivációs illetve emocionális állapotának reprezentációja, amely az előbbi mintájára nem szükségszerűen csak a pillanatnyi állapot felismerésére korlátozódik, hanem a korábbi tapasztalatok függvénye. E feldolgozási formának köszönhető, hogy bizonyos ágensek vonatkozásában megjelenhet például a barátság koncepciója, amely számos korábbi emocionális vonzatú (akár nem mindig szükségszerűen pozitív) reprezentációk eredője.
Az elmereprezentáció "hideg" aspektusának modellezéséhez Simon Baron-Cohen (1994) négy alapmodul létezését tételezi fel: (1) tekintetirány-érzékelés, (2) intencionalitás-érzékelés, (3) kölcsönös figyelem érzékelés, (4) másik elméjének reprezentációja. (A következőkben az angol mindreading mintájára a kissé körülményes "a másik elméjének reprezentációja" kifejezés helyett a tömörség kedvéért a magyarosabb "gondolatolvasási képesség" kifejezést azonos jelentésben használom.) A modulok közötti kapcsolat hierarchikus, hiszen az elme csak akkor képes a másik elméjének reprezentációjára, amennyiben rendelkezik a másik három modul által szolgáltatott információval. Érdemes megjegyezni, hogy a fenti modell nagyrészt humán (gyerek) vizsgálatok eredménye, és pillanatnyilag nincsen egyértelmű adat arra, hogy állatok esetében is működik az egész rendszer.
Idegi struktúrák szerepe a másik elmeállapotának reprezentációjában
Az utóbbi évek intenzív kutatásai nyomán számos, viszonylag jól körülírt idegi struktúráról derült ki, hogy szerepet játszik a másik elmeállapotának reprezentációjában (3. ábra).
Az idegrendszer moduláris felfogását előnyben részesítők számára a fő kérdés, hogy megtalálják-e a mentális képességhez elengedhetetlenül szükséges (obligát) struktúrákat, és megkülönböztessék azokat az alkalomszerűen, a feladat típusától függően bevont (fakultatív) struktúráktól (Siegel - Varley, 2002). E különbségtételre két módszer tűnik alkalmasnak. A disszociációs elv alapján csak az obligát struktúrák sérülése esetén figyelhető meg, hogy az elme teljes mértékben elveszti a megfelelő reprezentáció kiépítésére vonatkozó képességét, például amennyiben a nyelvi készség idegi struktúrái obligát részei a "gondolatolvasó" rendszernek, akkor ennek sérülése vagy funkciókiesése esetén az elme ezt a képességét is elveszti. Az ameliorációs módszer lényege, hogy csökkentjük a feladat olyan komponenseinek szerepét (beszéd, memória), amelyek elméleti szempontból nem lényegesek a "gondolatolvasási" képesség meglétéhez.
Sok kutató szerint a nyelvi képesség agyi struktúráinak fontos szerep jut a gondolatolvasó képességek megnyilvánulásában. Számos elemzés alapján azonban kiderült, hogy a speciális nyelvkészségbeli károsodást elszenvedők képesek megoldani gondolatolvasáson alapuló feladatokat, s a kutatók nem találtak összefüggést a károsodás mértéke, illetve a gondolatolvasási képesség sikeressége között egyedi szinten, ami arra utal, hogy bár ezen képesség megléte előnyös, de nem feltétlenül szükséges mások elmeállapotának reprezentációjához. Erre utal az is, hogy a már viszonylag komplex nyelvtannal beszélő hároméves gyerekek nem képesek gondolatolvasási képességet igénylő feladatok megoldására.
Egyes tapasztalatok szerint a frontális lebeny léziója esetén is csökken az ilyen feladatok megoldási sikeressége, s ehhez kapcsolódva említendő, hogy más struktúrák mellett épp a frontális lebeny mediális részén figyelhető meg agyi aktivitásnövekedés, miközben a kísérleti személyek ilyen feladatokat oldanak meg. Lézión átesett betegek esetében azonban nem találtak összefüggést a hagyományosan a frontális lebeny által kontrollált viselkedési akciókban (válogatás, gátlás, akciókövetés) való sikeresség, illetve a gondolatolvasási feladatokban való eredményesség között. Gondolatolvasási feladatok megoldása során aktivációt figyeltek meg a sulcus temporalis superiorban (STS) is. Az adott lézióval rendelkező betegeknek valóban nehezére esik az ilyen feladatok megoldása, mert nehezen választják szét a másik mentális állapotát a másik céljától. Sikerességüket azonban növelni lehetett úgy, hogy csökkentették a feladat olyan elemeit, amelyek "felesleges" terhet róttak a memóriára. Összességben úgy tűnik, hogy sem a frontális lebeny, sem a temporális-parietális kortex, nem tekinthető a gondolatolvasási képesség központi elemeinek.
A fent említett két módszer párhuzamos alkalmazásának első eredményei arra utalnak, hogy a gondolatolvasási képesség központi struktúráját az amigdala, illetve az azzal szoros kapcsolatban lévő mediális temporális lebeny jelenti. A bilaterális amigdala lézióval rendelkezők általánosságban igen rosszul teljesítenek gondolatolvasási feladatokban, és ezen a feladatok módosításával sem lehet segíteni. Az amigdala központi szerepe mellett szól az is, hogy ez a struktúra egyben a másik emocionális állapotának felismerésére is szolgál (lásd fent, az elmereprezentáció "meleg" aspektusa). Arcok látványa emberben és makákóban egyaránt aktiválja az amigdalát (különösen a félelmi gesztusok hatásosak). Bilaterális amigdala lézió barátságtalan viselkedés megjelenéséhez vezet, illetve az ilyen sérülést elszenvedő beteg elveszti hajlandóságát, hogy a fizikai (vagy absztrakt) világ eseményeit mentális terminusokban értelmezze.
Egyedfejlődési szempontok: tükrözés, kommunikáció, plaszticitás
A modell lehetséges komponenseinek felvázolása azonban önmagában még nem nyújt magyarázatot a modell működésére. A gondolatolvasási képesség modelljei általában két alapvető működési elvet tételeznek fel. A szimulációs elv szerint az egyed saját tapasztalatait terjeszti ki a másikra, azaz az elme megpróbálja elképzelni, hogy mit tenne a másik helyében, azaz a saját eszközeivel modellezi (szimulálja) a másik viselkedését. Ezen elképzelés ellen hangoztatott ellenérv, hogy akkor az elme nem lenne képes olyan mentális állapotot tulajdonítani a másiknak, amelyet ő maga nem is élt meg, azaz nem rendelkezik közvetlen tapasztalattal. Egy másik elképzelés szerint a gondolatolvasási képesség nem más, mint feltételezések (ún. teóriák) kialakítása a másik viselkedéséről, függetlenül a saját tapasztalattól. A két elképzelés közötti különbséget az alábbiak alapján szemléltethetjük: A szoba túloldalán látunk egy embert, aki velünk szembefordulva, előretekintve néz egy asztalon lévő tárgyat, előlünk azonban egy átlátszatlan paraván eltakarja az asztalt. A másik elmeállapotának elsődleges reprezentációját eszerint kétféle módon építhetjük fel. A szimulációs elv szerint elképzeljük, hogy a saját elménkben milyen reprezentáció jönne létre (az asztalra helyezett tárgy látványa), ha mi állnánk a szoba túloldalán. A "teóriaalkotó" elképzelés alapján, hogy a másik elmeállapota könnyen megjósolható, ha az elme rendelkezik a "látás" mint teória teóriájával, amely szerint az elmében "megjelenik" annak a tárgynak a képe, amelyet nézünk.
Giacomo Rizzolatti és munkatársai (1998) nemrég a temporális lebenyben (STS) olyan speciális neuronokat írtak le, amelyek az ágensek viselkedési értelmezésének egy új megközelítését teszik lehetővé. Az ún. tükörneuronoknak az az érdekessége, hogy nemcsak akkor tüzelnek, amikor az egyed egy bizonyos típusú akciót végző ágenst lát (lásd felismerő neuronok), hanem akkor is, amikor az egyed maga végzi az akciót. Így ezek a neuronok közvetlen kapcsolatot jelentenek a megfigyelő, illetve a szereplő viselkedése között. Ez a megfigyelés sokak szerint arra utal, hogy bizonyos esetekben az elme a szimuláció révén reprezentálja a másik akcióját (Gallese - Goldman, 1999).
Valójában azonban mindkét esetben szükség van előzetes tapasztalatokra, hiszen valószínű, hogy a látás mint teória felismerésére csak a szociális környezettel való bizonyos típusú interakciók nyomán vagyunk képesek. Az ilyen interakciók egyik fontos típusát képezi a kommunikáció. Pszichológusok régóta sejtik, hogy a másokkal történő verbális interakcióknak fontos szerepe lehet a gyerekek gondolatolvasási képességének kialakulásában. A feltételezés szerint megfelelő "beszélgetős" környezet kell ahhoz, hogy a gyerek értesüljön a másik mentális állapotáról, miközben a többi segédrendszerek érési folyamata zajlik. Négy-öt éves gyerekek biztosan oldanak meg gondolatolvasáson alapuló feladatokat, s lehet, hogy az idegrendszer érési folyamatait is figyelembe véve ennyi idő kell a megfelelő szociális tapasztalatok megszerzéséhez. Ezzel kapcsolatban érdekes megfigyelés, hogy azok a süket gyerekek, akik halló szüleik miatt csak későn tanulják meg a jelbeszédet, sokkal rosszabb gondolatolvasási képességgel rendelkeznek, mint a korábban süket szülőktől jelbeszédet elsajátító gyerekek, s ez a csökkent teljesítmény később már nem korrigálható. A jelenség magyarázható úgy, hogy az idegrendszer érésének érzékeny szakaszában csak ez utóbbiaknak van módjuk beszélgetni a szüleikkel.
Ember, csimpánz és kutya: homológiák és analógiák a szociális kogníció evolúciójában
A szociális kogníció vizsgálata jelenleg szinte kizárólag az emberről szól. A "hazai pálya" előnye természetesen érthető, amennyiben mind genetikai, mind neurobiológiai szempontból az ember talán a legjobban ismert faj, és a szociális kogníció folyamatai megértésének számos előnye van a gyógyítástól a nevelésen keresztül az emberi kultúra értelmezéséig. Két szempont azonban további megfontolást igényel. Legyen szó bármilyen biológiai jelenségről, az csak akkor értelmezhető a maga teljességében, ha tisztában vagyunk a kérdés evolúciós hátterével. Másfelől a természettudományokban bevett gyakorlat, hogy egyes komplexebb jelenségek értelmezése esetén a kutatók sokszor vizsgálnak olyan eseteket, amelyeknél a kérdéses jelenség egyszerűbb formában fordul elő. Ezeket szokás ún. állati modelleknek nevezni.
Az evolúció kérdése természetesen a szociális kognícióval kapcsolatban is felmerült. Az egyik széles körben népszerűsített elképzelés szerint a szociális kogníció megjelenésében és komplexitásának kialakulásában döntő szerepet játszott a szociális csoport, illetve a csoporttagok közötti interakció típusa. E nézet szerint az egyed akkor volt sikeres, ha társainál jobban volt képes megjósolni a másik várható akcióit, és ezt figyelembe venni saját akciójának tervezésekor. Különösen vonatkozik ez olyan komplex szociális helyzetekre, amikor az egyed érdeke, hogy akcióját a másik ne vegye észre, illetve amikor a másikat félre kell(ene) vezetnie. A mások ekképpen való "manipulálásának" képességét a híres egykor élt politikus nyomán szokás "machiavelliánus" intelligenciának is nevezni (Byrne - Whiten, 1988).
Az emberi szociális kogníció evolúciójának vizsgálatakor természetes választásnak tűnt más rokon főemlősök hasonló képességeinek vizsgálata. Mivel joggal feltételezhető, hogy a ma élő ember és csimpánz rendelkeznek közös ősökkel, sőt nagyobb időléptékben ugyanez mondható el az ember és az óvilági majmok viszonyáról, az evolúciós logika alapján feltételezhető, hogy az ember szociális képességeinek egy része meglehetősen ősi gyökerekkel rendelkezik, amelyeknek formáit a primáták esetében is megtaláljuk. Amikor a kutatók akár viselkedési, akár neurobiológiai szempontból vizsgálják a csimpánzok vagy makákók képességeit, feltételezik, hogy ezek evolúciós értelemben megfeleltethetők emberi tulajdonságokkal, más szóval homológiák.
Fontos azonban észrevenni, hogy a homológiák mentén való összehasonlítás csak az egyik lehetséges evolúciós megközelítés. Bár igaz, hogy az evolúciós rokonság miatt fennálló hasonlóságok összehasonlíthatók, de az is tény, hogy a közben eltelt idő alatt mind az ember, mind a vele rokon fajok is számos változáson mentek át, s nagy a valószínűsége, hogy már egyik faj sem él ugyanabban az ökológiai, illetve szociális környezetben, mint egykori közös ősei. Tehát bármennyire rokonnak tekinthetők az ember és a csimpánz, a szétválás óta eltelt idő elegendő lehetett arra, hogy olyan új szelekciós tényezők lépjenek fel, amelyek a közös eredeten alapuló hasonlóságot nagyrészt elmossák (gondoljunk csak arra, ránézésre ki gondolná, hogy a delfin uszonya és a mi végtagunk homológ szervek).
A fenti érvelést megfordítva viszont mondhatjuk, hogy hasonló környezetbe kerülve a fajok egyes tulajdonságai is hasonlíthatnak egymásra. Ilyenkor azonban nem a rokonság, hanem az adott szelekciós hatás vezet el a hasonló tulajdonságok megjelenéséhez. Az ilyen konvergenciák, illetve a szelekció következtében létrejött viselkedési analógiák vizsgálata kutatási szempontból éppúgy hatékony módszernek tekinthető, hiszen egy képesség megjelenése (akár több esetben, egymástól függetlenül is) talán még jobban segít megérteni e tulajdonság túlélés szempontjából fennálló jelentőségét.
Az ELTE Etológia Tanszékén kialakított kutatási irány épp egy ilyen alternatív módszer kidolgozását tűzte ki célul (lásd például Csányi - Miklósi, 1998; Miklósi, 2001; Miklósi et al., 2004). A munkánk alapját képező hipotézis szerint a csimpánz-ember homológia mintájára felvethető egy ember-kutya analógia, annak alapján, hogy az utóbbi 100-35 ezer évben a kutya ugyanabban az ökológiai és szociális környezetben élt. A kutya szempontjából nézve, feltételezzük, hogy azok az egyedek voltak képesek az ember közelében megmaradni, amelyek alkalmazkodtak az emberre jellemző viselkedési komplexumhoz (kötődés, kommunikáció, kooperáció stb., lásd Csányi, 2000). Ez a hosszú szelekciós folyamat vezethetett ahhoz, hogy a kutya szociális kognícióját tekintve megjelentek olyan analógnak tekinthető tulajdonságok, amelyek az emberéhez hasonlíthatók. Az utóbbi pár év kutatási eredményei egyértelműen igazolták ezt a feltételezést. A kutyák kötődési viselkedése, gesztusokon alapuló kommunikációja, szociális tanulási formái, illetve kooperációja funkcionális szempontból megfeleltethető az ember hasonló tulajdonságainak (Topál et al., 1998; Miklósi et al., 2000; Gácsi et al., 2001; Naderi et al., 2001; Pongrácz et al., 2001; Soproni et al., 2002).
A kutya mint az ember szociokognitív modellje különösen abban a tekintetben lehet jelentős, amikor a szociális tapasztalat hatását vizsgáljuk, hiszen a kutyák természetes környezetét az emberi közösség alkotja, ettől függetlenül egyedi szempontból tekintve igen eltérő lehet a szociális hatás, gondoljunk csak egy menhelyi kutya, illetve egy családi kedvenc közötti különbségre.
Jelen írás során a szerzőt az OTKA támogatta, további köszönet dr. Topál Józsefnek, valamint Csányi Vilmos professszornak, akinek lelkes támogatása nélkül e sorok talán sohasem születnek meg. További köszönet Topál Józsefnek kritikai megjegyzéseiért.
Kulcsszavak: szociális kogníció, reprezentáció, elmeteória, modularitás, tükrözés, kommunikáció, plaszticitás, evolúció
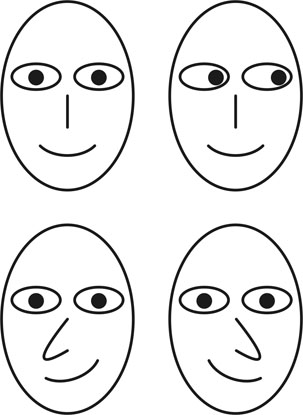
1. ábra * Tekintetfelismerő neuronjaink segítségével könnyen észleljük, hogy a felső sorban látható két stilizált fej merre tekint. A tekintet irányának észlelésében azonban a fej egyéb paraméterei is szerepet játszhatnak. Kísérleti alanyok (és talán az olvasó is) az alsó sorban lévő fejek esetében úgy érezték, hogy az illető a bal vagy a jobb oldaluk mellett néz el, annak ellenére, hogy a két képen a szemek pozíciója megegyezik.
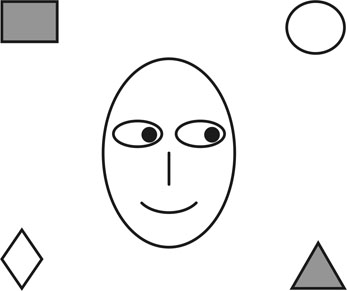
2. ábra * A képet nézők könnyen eldöntik, hogy az illető melyik tárgyat nézi. 2-3 éves gyerekek arra is tudnak válaszolni, hogy vajon az illető melyik "csokoládét" szereti. Autista gyerekek, akik számos teszt tanúsága szerint csökkent "gondolatolvasási" képességekkel rendelkeznek, ebben az esetben sem képesek ilyen következtetésre.
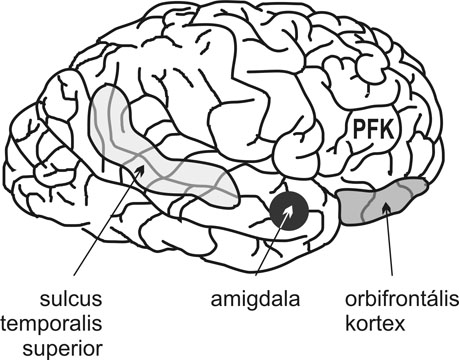
3. ábra * A gondolatolvasási képességben részt vevő idegi struktúrák elhelyezkedésének vázlatos ábrázolása az emberi agyban. PFK: prefrontális kéreg.
IRODALOM
Adolphs, Ralph (1999). Social Cognition and the Human Brain. Trends in Cognitive Sciences. 3, 469-79
Allison, Truett - Puce, Aina - McCarthy, Gregory (2000). Social Perception from Visual Cues: Role of the STS Region. Trends in Cognitive Sciences. 4, 267-77
Baron-Cohen, Simon (1994). How to Build a Baby That Can Read Minds: Cognitive Mechanisms in Mindreading. Current Psychology of Cognition. 13, 513-52
Brothers, Leslie - King, Brian (1992). A Neuroethological Framework for the Representation of Minds. Journal of Cognitive Neuroscience. 4, 107-18
Byrne, Richard W. - Whiten, Andrew (1988). Machiavellian Intelligence: Social Expertise and the Evolution of Intellect in Monkeys, Apes and Humans. Claredon Press, Oxford
Csányi Vilmos (2000). The 'Human Behaviour Complex' and the Compulsion of Communication: Key Factors of Human Evolution. Semiotica. 128, 45-60
Csányi Vilmos - Miklósi Ádám (1998). A kutya mint a korai emberi evolúció modellje. Magyar Tudomány. 9, 1043-54
Gácsi Márta - Topál J. - Miklósi Á. - Dóka A. - Csányi V. (2001). Attachment Behavior of Adult Dogs Living At Rescue Centers: Forming New Bonds. Journal of Comparative Psychology. 115, 423-31
Gallese, Vittorio - Goldman, Alvin (1999). Mirror Neurons and the Simulation Theory of Mind-Reading. Trends in Cognitive Sciences. 2, 493-500
Langton, Stephen R. H. - Watt, Roger J. - Bruce, Vicki (2000). Do the Eyes Have It? Cues to the Direction of Social Attention. Trends in Cognitive Sciences. 4, 51-58
Miklósi Ádám (2001). Állati modellek az emberi evolúció kutatásában. In: Pléh Csaba - Csányi Vilmos - Bereczkei Tamás (eds.): Lélek és evolúció. Osiris, Budapest, 97-118
Miklósi Ádám - Polgárdi R. - Topál J. - Csányi V. (2000). Intentional Behaviour in Dog-Human Communication: An Experimental Analysis of 'Showing' Behaviour in the Dog. Animal Cognition. 3, 159-66
Miklósi Ádám - Topál József - Csányi Vilmos (2004): Comparative Social Cognition: What Can Dogs Teach Us? Animal Behaviour. 67, 995-1004
Naderi Szima - Miklósi Á. - Dóka A. - Csányi V. (2001). Cooperative Interactions Between Blind Persons and Their Dog. Applied Animal Behaviour Science. 74, 59-80
Perrett, David I. - Smith, A. J. - Potter, D. D. - Mistlin, A. J. - Head, A. S. - Milner, A. D. - Jeeves, M. A. (1985). Visual Cells in the Temporal Cortex Sensitive to Face View and Gaze Direction. Proceedings of the Royal Society of London Series B. 223, 293-317
Pongrácz Péter - Miklósi Á. - Kubinyi E. - Gurobi K. - Csányi V. (2001). Social Learning in Dogs: The Effect of a Human Demonstrator on the Performance of Dogs (Canis Familiaris) in a Detour Task. Animal Behaviour. 62, 1109-1117
Povinelli, Daniel J. - Nelson, Kurt E. - Boysen, Sarah T. (1990). Inferences about Guessing and Knowing by Chimpanzees (Pan troglodytes). Journal of Comparative Psychology. 104, 203-10
Premack, David - Woodruff, Guy (1978). Does the Chimpanzee Have a Theory of Mind? The Behavioral and Brain Sciences. 1, 515-26
Rizzolatti, Giacomo - Arbib, Michael A. (1998). Langauge Within Our Grasp. Trends in Neurosciences. 12, 188-194
Siegal, Michael - Varley, Rosemary (2002). Neural Systems Involved in 'Theory of Mind'. Nature Reviews Neuroscience. 3, 463-71
Soproni Krisztina - Miklósi Á. - Topál J. - Csányi V. (2002). Comprehension of Human Pointing Gesture in Dogs. Journal of Comparative Psychology. 116, 27-34
Topál József - Miklósi Ádám - Csányi Vilmos (1998). Attachment Behaviour in the Dogs: A New Application of the Ainsworth's Strange Situation Test. Journal of Comparative Psychology. 112, 219-29