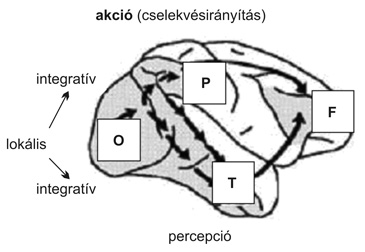
A látás fejlődése nem homogén folyamat, különböző funkciók különböző mértékben igénylik a tapasztalat befolyását, és eltérő sebességgel alakulnak ki. Két dimenzió mentén fogjuk elemezni a látás fejlődését (1. ábra): (1) Az integratív működés kialakulása, azaz a lokális-integratív dimenzió: a lokális ingertulajdonságok (például szín, orientáció, mozgás, mélység) mérése, s ezek téri integrációja. Míg a lokális folyamatok viszonylag korán kialakulnak, a téri integráció lassan fejlődik. A lokális-integratív dimenziót a hátsó nyakszirti agyterületekhez kötjük. (2) A két agykérgi látórendszer lassú kialakulása, s fejlődésbeli disszociációja, azaz az akció-percepció dimenzió. Ezen belül az occipitoparietális vagy a dorzális látórendszeri agyi struktúrák gyors fejlődése a látáson alapuló mozgásos (vizuomotoros) kontrollban kap szerepet; míg lassabb érés, fokozottabb hajlékonyság jellemezheti a tárgyak, események kategorizálását végrehajtó occipitotemporális vagy ventrális látórendszert.
1. Az integratív működés lassú kialakulása
A látás agykérgi folyamataival kapcsolatban ismereteink jelentős részét az állatokon végzett egysejt regisztrátumokat alkalmazó neurofiziológiai kísérletek adják. Ezek nyomán az az általános felfogás alakult ki, hogy az elsődleges látókéregbe érkező látási input egyszerű ingertulajdonságok (például orientáció, szín, luminencia, mozgás, retinális diszparitás) mentén kerül elemzésre, s ezt az elemzést viszonylag kis receptív mezővel rendelkező, tehát lokálisan működő neuronok hajtják végre. Pszichofizikai tanulmányok megerősítik ezt az elképzelést. A luminenciakontraszt által meghatározott élek, kontúrszegmensek elemzése esetén például adaptációs és maszkolási kísérletek során kiderült, hogy a retinakép elemzését több téri skálán működő, orientációhangolt szűrők végzik, melyek működése viszonylag lokális, tehát a látótérnek csak a szűrő méretével egyező darabjára vonatkozik (2. ábra). Nagy mennyiségű leíró jellegű adat gyűlt össze a lokális látási funkciók fejlődésével kapcsolatban az emberi élet első évében. Tudjuk például, hogy nagyon korai preferencia alakul ki a mozgó ingerekre (Nelson - Horowitz, 1987), két hónapos kortól kezdve jó a flickerérzékenység; két-három hónapos kor után megbízható a színérzékenység (Teller - Bornstein, 1986); négy hónapos korban megjelenik a retinális diszparitáson alapuló sztereopszis (Braddick et al., 1980) és a kontrasztérzékenység, s az ezáltal meghatározott látásélesség gyorsan növekszik az első életév során (Teller - Bornstein, 1986).
A kontrasztérzékenységet meghatározó téri szűrők korai érési mintázatát csecsemők esetén az ún. nézéspreferencia technikával lehet tanulmányozni (például Teller - Bornstein, 1986). A használt ingerek általában szinuszos luminenciaeloszlású rácsmintázatok (3. ábra). Egy adott téri frekvenciájú, orientációjú és kontrasztú mintázatot párban mutatnak be egy homogén szürke képpel, és regisztrálják, hogy melyik mintázatot nézi a csecsemő hosszabb ideig. Amennyiben a csecsemő még nem képes az adott frekvenciájú rácsot vizuálisan felbontani, mindkét kép egyformán homogén szürkének fog látszani számára, és nagyjából a nézési idő 50-50 %-át fogja tölteni velük. Ha már látja az adott rácsmintázatot, akkor ezt a számára érdekesebb képet, a nézési idő több mint 50 %-ában fogja szemlélni. A kontraszt változtatásával azután meg lehet mérni, hogy mi a csecsemő látási küszöbe egy adott téri frekvenciájú rácsra. A küszöböt általában a 75 %-os nézési idővel definiálják, és számos téri frekvencián meghatározzák. Ezen mérések alapján objektíven meg lehet mérni, hogy mi is a csecsemő rendelkezésére álló információ a látható fény luminenciakontraszt által meghatározott tartományában, és meg lehet rajzolni a csecsemő kontrasztérzékenységi görbéjét. Ez a görbe nemcsak laposabb lesz, mint a felnőtté, de a csúcsa az alacsonyabb téri frekvenciák felé tolódik el. Ez azt jelenti, hogy a csecsemő csak a tárgyak durva részleteit látja. A fenti nézéspreferencia-vizsgálatot a többi, már említett ingertulajdonság esetén is el lehet végezni. A 4. ábra luminenciakontraszt-érzékenységi méréseken alapuló illusztráció, mely azt hivatott bemutatni, hogy miként alakul az érzékenység az élet első hat hónapjában. Az illusztrált fejlődés nagyrészt a fent említett lokális elemző folyamatok érése révén valósul meg.
Amint egy bizonyos funkció megjelenésének idejét megbecsültük, az ezt követő fejlődés a következő kérdés. Ez azonban módszertanilag sokkal bonyolultabb, mert összehasonlító és nem abszolút (egy bizonyos funkció jelen van-e avagy nem) viselkedési vagy fiziológiai méréseken alapszik. Egy bizonyos feladatban nyújtott teljesítményt össze kell hasonlítani egy idősebb vagy felnőtt csoporttal. A nehézségek egyike, hogy a teljesítmény szintjét befolyásolják nem-vizuális tényezők, mint a motiváció vagy a figyelem hiánya, és az adatok összehasonlíthatósága a felnőtt csoportokéval emiatt nem mindig egyértelmű. Ez lehet az egyik ok, amiért az emberi látásfejlődés viselkedéses vizsgálatai a második év után nem túl számosak, bár az elmúlt évtizedben volt példa néhány jól kontrollált vizsgálatra. Újabb vizsgálatok kiemelik az ingerek időbeli jellegzetességeit a fejlődési görbék meghatározásánál. A látásélességen túlmenően úgy tűnik, hogy a textúrán, mozgáson és színen (Hollants-Gilhuijs, Ruijter és Spekreijse, 1998) alapuló vizuális szegmentáció és a formaazonosítás viszonylag lassan kialakuló, második életév után beérő képességek. Pontosan mely mechanizmusok húzódnak a gyermekek rossz teljesítményének hátterében ezeknél a feladatoknál? Úgy tűnik, mindegyik feladat magában foglal a vizuális tulajdonságok lokális elemzésén túli mechanizmusokat is, és mindegyik a látómező egészén vagy nagyobb darabján megjelenő információ integrációjának képességén alapszik.
A teljes látómezőben vagy a látómező nagyobb szegmensén belül megjelenő információ integrálása a tulajdonságok lokális elemzésén túlmenő folyamatokat igényel. Kiragadva egyet az elsődleges ingertulajdonságok közül, az orientáció információ integrálásáról fogok beszélni. A lokális orientációmérések integrációja az alacsony szintű feldolgozás egyik igen fontos mechanizmusa, mely a tárgyak határait alkotó kontúrok kiemelését teszi lehetővé. Ez egy olyan köztes feldolgozási szint, amely még messze van a tárgyazonosítástól, de már implikálja a tárgyak háttértől való elkülönítését. Neurális háttere az elsődleges látókéreg orientációhangolt neuronjaik szintén orientációhangolt interakcióin, axonális összeköttetésein alapul (Gilbert, 1998). Pszichofizikailag egy ún. kontúrintegrációs paradigmán belül lehet vizsgálni (például Field, Hayes és Hess, 1993; Kovács és Julesz, 1993).
A kontúrszakaszok egybekötéséért felelős képességünket vezérlő neurális kapcsolatok szabályait egy zajba ágyazott Gábor-foltokból folyamatos ívet alkotó ingeregyüttessel vizsgáljuk (5. ábra). A Gábor-foltok modellezik az elsődleges látókéregben lévő irányérzékeny sejtek receptív mezejének tulajdonságait, és így optimális ingert nyújtanak számukra. A beágyazott kontúrokat nem észlelhetjük kizárólag lokális szűrőkkel, vagy az ismert irányérzékeny, nagy receptív mezővel rendelkező neuronokkal. A hosszú távú orientációs együtt járások észlelése a kontúr nyomvonala mentén csak a lokális mérések integrációjával lehetséges. A zaj arra kényszeríti a megfigyelőt, hogy ezeket a méréseket az egyes Gábor-foltok szintjén végezze, és kizárólag a lokális szűrők hosszú távú kapcsolataira támaszkodjon a foltok perceptuális összekötése során. Így ezekkel az ingerekkel a téri integrációt szolgáló hosszú távú kapcsolatokat elkülönítve vizsgálhatjuk. Ezek a viszonylag alacsony szintű interakciók érzékenynek tűnnek a perceptuális csoportosítás tényezőire. Ebben a feladatban a zárt kontúrok fölényét mutattuk ki a nyílt kontúrokkal szemben a szomszédos elemek maximális távolságának szempontjából (Kovács - Julesz, 1993), és fokozott lokális kontrasztérzékenységet a zárt kontúrokon belüli mezőben (Kovács - Julesz 1994). Ezen interakciók megvalósulási szintjén egy tárgyközpontú formareprezentáció létezésére utaló jeleket is találtunk (Kovács - Julesz, 1994; Kovács, 1996; Kovács et al., 1998). A fenti eredmények az interakciók tárgyakhoz kapcsolódó feldolgozásban való részvételét jelzik.
Az irányultsági információ téri integrálását a kontúrdetekciós feladatban (Field et al., 1993; Kovács - Julesz, 1993; Kovács, 1996) és egy kevésbé bonyolult laterális maszkolási paradigmát alkalmazó konfigurációban (Polat - Sagi, 1994) az irányultságra hangolt téri csatornák hosszú távú serkentő jellegű interakcióihoz kapcsolták. A pszihofizikailag leírt hosszú távú interakciók lehetséges anatómiai alapját az elsődleges vizuális kérgen belüli horizonális kapcsolatokat alkotó axonok képezik (Gilbert, 1998). A kontúr feladathoz hozzájárulhatnak még az extrastriatális kéregből eredő visszacsatolást biztosító összeköttetések, amik a felülről lefelé irányuló folyamatokért felelősek. Egy nemrégiben végzett neuropszichológiai vizsgálatban (Giersch et al., 2000), egy látási agnóziás, sértetlen V1-gyel rendelkező kísérleti személy jól teljesített a kontúrintegrációs feladatban, annak ellenére, hogy a magasabbrendű occipitális kérgi területei súlyosan roncsolva voltak. Ez az eredmény arra utal, hogy az elsődleges vizuális kéreg önmagában elegendő lehet a kontúrintegrációhoz.
A kontúrintegrációs feladat egyszerűsített változatát használtuk fel a vizuális téri integráció fejlődésének vizsgálatára. Gyerekeknél öt és tizennégy év között jelentős teljesítményjavulást (Kovács et al., 1999), korai éveikben rendellenes binokuláris ingerlésnek kitett felnőtteknél pedig rossz teljesítményt találtunk (Kovács et al., 1999). Ezenkívül nagyon fiatal (három hónapos csecsemőket, Gerhardstein et al, 2004), illetve különböző betegségben szenvedő, feltehetően perceptuális organizációs problémával küszködő egyéneket (Giersch et al., 2000; Silverstein et al., 2000) is vizsgáltunk. Ezekben a vizsgálatokban a kontúrdetekciós feladatot kártyákon mutattuk be (Kovács et al., 1999, 2000). A kártyaváltozat jobban alkalmazható gyermekeknél és klinikai környezetben, és egyben több kísérleti személy vizsgálatára ad lehetőséget. Az 5. ábra illusztrálja, hogy a kontúrintegrációs kártya egy Gábor-foltokból képzett zárt kört (kontúr), valamint véletlenszerű irányultságú és pozíciójú Gábor-foltokat (orientációs zaj) tartalmaz. A feladat a kontúr nyomvonalának követése. Annak érdekében, hogy a hosszú távú interakciók erejét meg tudjuk becsülni egy adott egyén esetén, a háttérzaj sűrűségét változtattuk. A relatív zajsűrűséget (D) a zajelemek átlagos távolságának és a kontúrelemek távolságának arányaként határozzuk meg (5. ábra). Ha D < 1, a kontúrt az elemek sűrűségi információja alapján is észlelhetjük, mert a kontúr elemei közelebb vannak egymáshoz, mint a zaj elemei. Ha azonban D > 1, a sűrűség nem jelent segítséget, és a kontúrt lehetetlen orientációhangolt, hosszú távú interakciók nélkül azonosítani. A D küszöbértéke (Dmin, azaz a D értéke a legnehezebb azonosított kártyán) meghatározza ezen interakciók erősségét az egyes kísérleti személyekben. Minél kisebb ez az érték, annál erősebbek az interakciók.
Több mint négyszáz normális látású, öt és tizennégy év közötti gyereket vizsgáltunk a kártyás eljárással (Kovács et al., 1999). Amint azt a 2. ábrán láthatjuk, a tizenhárom és tizennégy év közötti gyerekek a kontúrok többségét jól azonosították a kártyasorozatban (Dmin=0,7), míg az öt és hat év közötti gyerekek a kártyák felénél átsiklottak a kontúrok fölött. Ez igen nagy különbség kontúrintegrációs teljesítményben a két csoport között. A teljesítmény fokozatosan emelkedik, és a felnőtt szintet csak valamikor a serdülőkor után éri el ebben a feladatban. Felmerült az a kérdés, hogy ez az igen meglepő eredmény és a nagyon lassú fejlődés valóban perceptuális fejlődésnek tulajdonítható, vagy figyelmi, motivációs folyamatok állnak-e inkább a héttérben. Ennek eldöntésére egy perceptuális tanulási kísérletet végeztünk, melyben megvizsgáltuk, hogy egyrészt van-e tanulás ebben a feladatban, másrészt mennyire specifikus a tanulás az adott ingertulajdonságokra (Kovács et al., 1999). Tudni kell azt, hogy az alacsony szintű perceptuális tanulás jellegzetessége az ingerspecifikusság. Tehát ha például javulást érünk el egy olyan feladatban, ahol orientációjelzések határozzák meg a detektálandó ingert, a javulás nem fog áttevődni a feladat azon változatára, ahol az orientációt színnel vagy más jelzésekkel helyettesítettük (Fiorentini - Berardi, 1980, Karni - Sagi, 1993). A nagyfokú ingerspecificitás általában azt jelzi, hogy a tanulás plasztikus neuronális változásai alacsony szintű kérgi területeken játszódtak le, ahol az ingerdimenziók még elkülöníthetőek. Ez a fajta specifikusság nem áll fenn figyelmi vagy motivációs tanulásnál. Kísérletünkben egy új kártyasorozatot vezettünk be, amelyben a kontúr íve színnel, és nem orientációval volt jelezve. A szín által meghatározott kártyák nehézségi foka megegyezett az eredeti kártyák nehézségi fokával. Pár nap gyakorlás után jelentős, orientáció, illetve színspecifikus tanulást figyeltünk meg gyerekeknél és felnőtteknél egyaránt. Úgy tűnik, hogy a kontúrészlelési feladatban a tanulás eredménye nem átvihető az irányultságról a színre, ez pedig azt jelzi, hogy a háttérben álló mechanizmusok specifikusan hangolt észlelési mechanizmusok.
Miért teljesítenek a gyerekek rosszul a kontúrintegrációs feladatban? Az elsődleges látókéreg két-három rétegének vízszintes összeköttetéseiről kimutatták, hogy fejlődésük jócskán belenyúlik a gyermekkorba (Burkhalter, 1993), sőt emberekben a V1 és V2 közötti visszacsatoló összeköttetések késleltetett, születés utáni fejlődésére is vannak jelek (Burkhalter, 1993). További kísérletet végeztünk, hogy megállapítsuk, vajon az abszolút kérgi távolságokat áthidaló laterális interakciók hossza korlátozza-e a kontúrintegrációs feladatban nyújtott teljesítményt. A kísérletben a kontúr elemei közt a távolságot változtattuk, de a viszonylagos zajszintet állandónak hagytuk. Azt találtuk, hogy a Dmin felnőttekben független a kontúrt alkotó elemek távolságától. Ez arra utal, hogy a felnőtteket csak a képi paraméterek (jel-zaj arány) korlátozzák a vizsgált távolságtartományban, a kérgi interakciók abszolút távolsága viszont nem. Gyerekeknél azonban a kontúrelemek közötti kisebb távolságnál jobb volt a teljesítmény, és a gyermekek és felnőttek közötti különbségek a kontúrészlelésben nagyobb távolságoknál még nagyobbak voltak (Kovács et al., 1999). Arra következtettünk, hogy gyerekeknél a hosszú távú interakciók rövidebb téri távolságot hidalnak át, mint felnőtteknél.
Mivel a kontúrteszttel vizsgált interakciók általánosabb értelemben nemcsak a kontúrok, hanem a vizuális kontextus integrációját is szolgálják, további vizsgálatokat végeztünk arra nézve, hogy milyen következményei lehetnek a hosszú távú neurális kapcsolatok éretlenségének a gyerekek "perceptuális világával" kapcsolatban. Így például egy geometriai vizuális illúziót, az ún. Ebbinghaus-illúziót (6. ábra) felhasználva megvizsgáltuk, hogy van-e eltérés a felnőtt és a négyéves gyermek illúzióbenyomásában (Káldy - Kovács, 2003). Az Ebbinghaus-illúzió esetén a két középső kör egy-egy lokális ingernek tekinthető, melyeket a kontextusként elhelyezett kis, illetve nagy körök akkor tudnak befolyásolni, ha az összes ingerelem téri integrációja megtörténik a látómezőn keresztül. Míg felnőttek esetén az illúzió mértéke nagyon világosan meghatározható, s mindig ugyanabba az irányba mutat (a nagy körök által körülvett kör tűnik kisebbnek), négyéves gyerekek általában nem tapasztalnak illúziót, vagy ha igen, az nagyon kis mértékű és bizonytalan irányú (Káldy - Kovács, 2003). Egy másik esetben a binokuláris rivalizáció példáját vizsgáltuk meg, ahol a két szem számára mesterségesen, egészen eltérő képeket mutatunk be (Kovács - Eisenberg, 2004). Ebben a kísérletben az egyik szem függőleges, a másik vízszintes rácsmintát szemlél, s a felnőtt megfigyelő a két mintázat rivalizálásából származó spontán váltakozást figyelhet meg a vízszintes és függőleges rácsminták között. Négy-öt éves gyerekek azonban úgy tűnik, vagy egyáltalán nem látnak váltakozást (tehát például csak vízszintes mintát látnak), vagy a két mintázat apró foltjainak együttes jelenlétét észlelik ebben az esetben (Kovács - Eisenberg, 2004). Ezt az eredményt ismét az integráció hiányával hoztuk összefüggésbe. Végül, a harmadik érdekesség, amit szeretnék említeni, szintén egy klasszikus érzékelési demonstrációhoz kötődik, az ún. "foltos" képekhez. A 7. ábra felső ábráját a legtöbb felnőtt megfigyelő csak akkor ismeri fel, ha az alsó ábrán szereplő segítő rajzot is bemutatjuk, ekkor azonban már mindenki könnyedén meglátja a foltokban az arcot. Nem így az öt év körüli gyerekek, akik még akkor sem képesek az arcot felfedezni, ha a két képet egyszerre mutatjuk meg nekik. Ezt a megfigyelést a fixáció képen belüli mintázatának követését felhasználó kísérletben is megerősítettük. Azt találtuk, hogy míg a felnőttek foltos képekre adott, eredetileg random fixációs mintázata átrendeződik a segítő inger bemutatása után, addig a gyerekeké random marad (Kovács - Kovács, 2004). A foltok összeillesztéséhez feltehetőleg érett neurális interakciók szükségesek mind az elsődleges látókérgen, mind a magasabb látóterületeken belül.
Összegezve az eddigieket, úgy tűnik, hogy a lokális vizuális tulajdonságokat feldolgozó neurális elemek érnek először, míg a lokális tulajdonságokat koherens egésszé integráló hálózatok később szilárdulnak meg.
2. A két agykérgi látórendszer lassú kialakulása, s fejlődésbeli disszociációja
A majmokat leíró élettani, anatómiai és a humán pszichofiziológiai eredményekből régóta tudjuk, hogy a látásnak legalább két szakosodott alrendszere létezik. Korábbi beszámolók szerint a dorzális (occipitoparietális) pálya a térlátásra, míg a ventrális (occipitotemporális) pálya a tárgyfelismerésre szakosodott (Ungerleider - Mishkin, 1982). David A. Milner és Melvyn A. Goodale (1992) új értelmezésben szemlélik a funkcionális szétválást: a két pálya eltérő műveleteket hajt végre a mindkettőjük által hozzáférhető információn a tárggyal és annak téri elhelyezkedésével kapcsolatban. A dorzális pálya feldolgozása közvetíti a vizuálisan irányított mozgás kontrollját, míg a ventrális pálya lehetővé teszi az észlelt tárgy képi és kognitív reprezentációjának és jelentőségének kialakítását (Milner - Goodale, 1992). A korábbi megközelítések (Ungerleider - Mishkin, 1982) a két pálya közti képi attribútumok (mint a mozgás és téri elhelyezkedés a dorzális, szemben az irányultság, szín, méret és forma a ventrális látópályában) feldolgozásának munkamegosztására összpontosítottak, és azt feltételezték, hogy a két pálya kimenete a világ cselekvést és tudatos észlelést egyaránt szolgáló egységes reprezentációjában egyesül. Goodale és Milner kisebb hangsúlyt fektetnek a képi attribútumokat feldolgozó párhuzamos folyamatokra, inkább a filogenetikailag releváns funkcionális szempontokra összpontosítanak. Feltételezésük szerint a két párhuzamos látópálya kialakulásának oka a mozgás/cselekvés eltérő reprezentációs igénye a filogenetikailag sokkal fiatalabb képi percepcióval szemben. A reprezentációkhoz a két látópálya által végzett különböző műveleteken keresztül férhetünk hozzá. A cselekvő rendszer feladata a megfigyelőhöz képest pillanatról pillanatra változó tárgyak tulajdonságainak kódolása, ennek megfelelően ez a rendszer on-line, egocentrikus koordinátákat használ, és igen kicsi a memóriaterjedelme. Az észlelő rendszer feladata viszont a tárgyak azonosítása a kognitív rendszer általi további feldolgozás érdekében, ezért ez a pálya tárgyközpontú leírásokra és hosszú távú raktározásra alapoz. A két pálya kimenete két párhuzamos képi világot hoz létre, az egyik - főképp tudattalan - a cselekvés irányítását végzi, a másik a tárgyak állandó tulajdonságainak tudatos képi tapasztalatával lát el minket.
Amennyiben a két látópálya a bejövő információn célirányos cselekvés és észlelés érdekében végzett, valóban eltérő műveletek végrehajtására fejlődött ki, bármilyen fejlődési különválást specifikusan azt a pályát, dimenziót megcélzó feladatok segítségével fedhetünk fel. Az eszményi vizsgálat szelektíven a ventrális vagy dorzális látópályát igénybe vevő pszichofiziológiai feladatokat tartalmazna, és egyben azok fejlődési sebességéről, almintázatairól és lehetséges szétválásairól nyerne adatokat. Szelektíven az egyik pályát célzó feladatokkal a ventrális és dorzális pályák fejlődési görbéit össze tudnánk hasonlítani egymással. Ezt azonban igen nehéz elérni, mert a legtöbb létező pszichofiziológiai eljárás a ventrális pálya funkcióihoz kötődik. Vegyük észre, hogy az elméleti keret szerint (Milner -Goodale, 1992), a funkcionális különválás túlmutat a "hol" vs. "mi" (Ungerleider - Mishkin, 1982) vagy a "mozgás" vs. "azonosítás" (Braddick et al., 2003) típusú szétválasztáson. A két pálya jellegzetes tulajdonságai a "cselekvés" vs. "észlelés" dimenziók mentén helyezkednek el, és a végső cél ezen dimenziók vizsgálata.
Hogyan tudjuk a ventrális pálya funkcióját a legjobban vizsgálni? "A ventrális pálya által végzett műveletek lehetővé teszik a tárgyak állandó tulajdonságait és a tárgyak viszonyait magukban foglaló perceptuális reprezentációk alkotását" (Goodale, 1998). A pályához tartozó leginkább releváns kérgi terület az inferotemporális (IT) kéreg. Az IT-t fontosnak tartjuk a tárgyállandóság, a nagyság és helykonstancia, valamint a rövid távú memória fenntartásában, és alapos tanulmányozásnak lett alávetve viselkedéses és kombinált, léziós vizsgálatokban késleltetett mintapárosítási (delayed matching-to-sample - DMS) feladat segítségével. Gyermekeknél a korlátozott távolságú téri perceptuális és csökkent kontextuális integrációval kapcsolatos eredményeinkre (Kovács et al., 1999) alapozva feltételezhetjük, hogy a DMS feladatban fontos perceptuális reprezentációk sokkal gyengébbek fiatal egyéneknél. Valóban, makákó majmoknál két további érési év szükséges a feladat egy változatának (delayed nonmatching-to-sample - DNMS) felnőtt szintű megoldásához (Bachevalier, 1990). Fény derült arra is, hogy még harminckét hónapos embercsecsemőknek is meghosszabbított tanulásra és érésre volt szükségük a DNMS feladat sikeres megoldásához (Overman et al., 1992).
Miként tudjuk a dorzális pálya funkcióját a legcélravezetőbben vizsgálni? A dorzális pálya "énközpontú referenciakeretet, a tárgyak egyik pillanatról a másikra hozzáférhető információit használja fel" (Goodale, 1998). Egy új keletű nézet szerint a posterior parietális kéreg (PPC) funkciója a szenzoros információ integrálása a cselekvéstervezéshez (Andersen - Bueno, 2002). A posterior parietális kéregben a látványelemek elhelyezkedése több koordinátarendszer szempontjából is kódolt, mint például szem, kéz, test és világ. A többi modalitásból származó szenzoros bemenetek itt keverednek, integrálódnak a vizuális jelekkel, hogy megfelelő motoros koordinátákkal szolgáljanak a célirányos mozgások kivitelezéséhez (Andersen - Bueno, 2002). A hatékony motoros koordinációhoz valószínűleg szükség van tanulási és érési folyamatokra is ezekben a struktúrákban, sajnos ezek fejlődésének természete és idői lefolyása azonban még nem ismert.
Vajon a párhuzamos pályák azonos ütemben fejlődnek? Feltevésünk szerint a percepciót szolgáló ventrális pálya késleltetve érik a cselekvést segítő dorzális pályához képest. A feltételezést a következő megfontolásokra alapozzuk: (a) makákókon végzett anatómiai megfigyelések az occipitoparietálishoz képest az occipitotemporális pálya lassabb funkcionális érését jelzik (Distler et al., 1996); (b) emberi anatómiai adatok a ventrális pálya kapcsolatainak a dorzális pályához viszonyított lassabb kialakulását mutatják (Kovács et al., 1999); (c) a dorzális pálya filogenetikailag ősibb, ami erőteljesebb genetikai kódoltságra és gyorsabb érésre enged következtetni; (d) az ontogenezis (egyedfejlődés) során nagyobb szükség lehet a cselekvés vizuális irányításáért felelős agyi képletek (dorzális pálya) korai kifejlődésére, mint a tárgyak felismeréséért felelős struktúrákéra (ventrális pálya); (e) ha a ventrális pálya valóban a környezetünk megértését elősegítő magas szintű kognitív hálózat részét alkotó perceptuális reprezentációk kialakításáért felelős, akkor ennek elnyújtott fejlődése és ezzel a korai időszakon túli plaszticitás megőrzése kívánatos.
Bár az első életév során már sok fontos látási funkció kialakul, a látás fejlődése, úgy tűnik, eltart egészen a gyermekkor végéig. A különböző látási funkciók, és a látási információ feldolgozásában részt vevő neuroanatómiai hálózatok kialakulása nem homogén folyamat. Két releváns dimenziót fedtünk fel, amelyekben a látás fejlődési mintázatait vizsgálhatjuk. A lokális-integratív dimenzió tanulmányozására kontúrintegrációs feladatot alkalmaztunk. Az adatok szerint a látómezőben lévő orientációs információ integrálásáért felelős neurális kapcsolatok rövidebb távolságot hidalnak át gyerekeknél, és csak a gyermekkor vége felé válnak a felnőttekéhez hasonlóvá. Továbbá feltételezzük, hogy viszonylag fejlett vizuomotoros koordináció mellett a ventrális pálya által végrehajtott látási funkciók kifejlődése szintén lassú fejlődésmenetet mutat (friss összefoglaló monográfia erről: Kovács, 2004).
Kulcsszavak: látás, fejlődés, kontraszt, kontúrintegráció, dorzális pálya, ventrális pálya
1 A cikk részleteket tartalmaz a következő munkából: Kovács Ilona (2000): Human Development of Perceptual Organization. Vision Research, 40, 1301-1310. Fordította: Gábris Krisztián.
IRODALOM
Andersen, Richard A. - Buneo, Christopher A. (2002): Intentional Maps in Posterior Parietal Cortex. Annual Review of Neuroscience. 25, 189-220.
Bachevalier, Jocelyne (1990): Ontogenetic Development of Habit and Memory Formation in Primates. In: Diamond Adele (ed.): Development and Neural Bases of Higher Cognitive Functions. Academic Press, New York
Braddick, O. J. - Atkinson, J. - Julesz B. - Kropfl, W. - Bodis-Wollner, I. - Raab, E. (1980): Cortical Binocularity in Infants. Nature. 288, 363-365.
Braddick, Oliver - Atkinson, J. - Wattam-Bell, J. (2003): Normal and Anomalous Development of Visual Motion Processing: Motion Coherence and 'Dorsal-Stream Vulnerability'. Neuropsychologia. 41, 13, 1769-1784.
Burkhalter, Andreas (1993): Development of Forward and Feedback Connections between Areas V1 and V2 of Human Visual Cortex. Cerebral Cortex. 3, 5, 476-487.
Distler, Claudia - Bachevalier, J. - Kennedy, C. - Mishkin, M. - Ungerleider, L. G. (1996): Functional Development of the Corticocortical Pathway for Motion Analysis in the Macaque Monkey: A 14C-2-Deoxyglucose Study. Cerebral Cortex. 6, 2, 184-195.
Field, David J. - Hayes, A. - Hess, R. F. (1993): Contour Integration by the Human Visual System: Evidence for a Local “Association Field". Vision Research. 33, 2, 173-193.
Fiorentini, Adriana - Berardi, Nicoletta (1980): Perceptual Learning Specific for Orientation and Spatial Frequency. Nature. 287, 43-44.
Gerhardstein, Peter - Kovács I. - Ditre J. - Fehér Á. (2004): Detection of Contour Continuity and Closure in 3-Month-Old Human Infants. Vision Research, 44, 2881-2988.
Gilbert, Charles D. (1998): Adult Cortical Dynamics. Physiological Review. 78, 467-485. http://www.its.caltech.edu/~kai/cortical_dynamics.pdf
Giersch, Anne - Humphreys, G. - Boucart, M. - Kovács I. (2000): The Computation of Occluded Contours in Visual Agnosia: Evidence for Early Computation Prior to Shape Binding and Figure-Ground Coding. Cognitive Neuropsychology. 17, 8, 731-759.
Goodale Melvyn, A. (1998): Vision for Perception and Vision for Action in the Primate Brain. Novartis Foundation Symposium. 218, 21-34.
Hollants-Gilhuijs, Marjorie A. M. - Ruijter, J. M. - Spekreijse, H. (1998): Visual Half-Field Development in Children: Detection of Colour-Contrast-Defined Forms. Vision Research. 38, 5, 645-649.
Káldy Zsuzsa - Kovács Ilona (2003): Visual Context Integration Is Not Fully Developed in 4-Year-Old Children. Perception. 32, 6, 657-666.
Karni, Avi - Sagi, Dov (1993): The Time Course of Learning a Perceptual Skill. Nature. 365, 250-252.
Kovács Ilona (1996): Gestalten of Today: Early Processing of Visual Contours and Surfaces. Behavioural Brain Research. 82, 1, 1-11.
Kovács Ilona (ed.) (2004): Visual Integration: Development and Impairments. Akadémiai, Budapest
Kovács Ilona - Eisenberg, Michal (2004): Human Development of Binocular Rivalry. In: Alais, David - Blake, Randolph (eds.): Binocular Rivalry. MIT Press, 100-116.
Kovács Ilona - Fehér A. - Julesz B. (1998): Medial-Point Description of Shape: A Representation for Action Coding and Its Psychophysical Correlates. Vision Research. 38, 15-16, 2323-2333.
Kovács Ilona - Julesz Béla (1993): A Closed Curve Is Much More Than an Incomplete One: Effect of Closure in Figure-Ground Segmentation. Proceedings of the National Academy of Sciences of the USA. 90, 16, 7495-7497.
Kovács Ilona - Julesz Béla (1994): Perceptual Sensitivity Maps within Globally Defined Visual Shapes. Nature. 370, 6491, 644-646.
Kovács Ilona - Kovács Gyula (2004): Lack of "One-Shot" Learning in Preschool Children (Eye-Movement Data. Vision Sciences Society Fourth Annual Meeting, Sarasota, Florida, 36.
Kovács Ilona - Kozma P. - Fehér A. - Benedek G (1999): Late Maturation of Visual Spatial Integration in Humans. Proceedings of the National Academy of Sciences of the USA. 96, 21, 12204-12209.
Kovács Ilona - Polat, U. - Pennefather, P, M. - Chandna, A, - Norcia, A. M. (2000): A New Test of Contour Integration Deficits in Strabismus and Amblyopia. Vision Research, 40, 13, 1775-1783,
Milner, A. David - Goodale, Melvyn A. (1995): The Visual Brain in Action. Oxford University Press, Oxford
Nelson, Charles A - Horowitz, Frances Degen (1987): Visual Motion Perception in Infancy: A Review and Synthesis. In: Salapatek, Philip - Cohen, Leslie (eds.): Handbook of Infant Perception 2. Academic, New York, 123-153.
Overman, William - Bachevalier, J. - Turner, M. - Peuster, A. (1992): Object Recognition Versus Object Discrimination: Comparison between Human Infants and Infant Monkeys. Behavioral Neuroscience. 106, 1, 15-29.
Polat, Uri - Sagi, Dov (1994): The Architecture of Perceptual Spatial Interactions. Vision Research. Jan, 34, 1, 73-78.
Silverstein, Steven - Kovács I. - Corry, R. - Valone, C. (2000): Perceptual Organization, the Disorganization Syndrome, and Context Processing in Chronic Schrizophrenia. Schizophrenia Research. 43, 1, 11-20.
Teller, David Y. - Bornstein, Marc H (1986): Infant Color Vision and Color Perception. In: Salapatek, Philip - Cohen, Leslie B. (eds.): Handbook of Infant Perception. Academic Press, New York
Ungerleider, Leslie G. - Mishkin, Mortimer (1982): Two Cortical Visual Sytems. In: Ingle, David J. - Goodale M. A. - Mansfield, R. J. W. (eds.): Analysis of Visual Behavior. MIT Press, Cambridge, MA, 549-586.