Vélemény, vita
A
MON 810-es GM-kukoricák
környezettudományi
megítélése
|
Darvas
Béla |
Lauber
Éva |
|
az
MTA doktora, c. egyetemi tanár |
tudományos
segédmunkatárs |
|
MTA
Növényvédelmi Kutatóintézete, |
MTA
Növényvédelmi Kutatóintézete, |
|
Ökotoxikológiai
és Környezetanalitikai O., Bp. |
Ökotoxikológiai
és Környezetanalitikai O., Bp. |
|
bdarvas
 freemail.hu
freemail.hu |
eva.lauber
gmail.com |
|
|
|
|
Bakonyi
Gábor |
Békési
László |
|
az
MTA doktora, egyetemi tanár |
az
állatorvos-tud. kandidátusa, c. egyetemi tanár |
|
Szent
István Egyetem, |
Állattenyésztési
és Takarmányozási Kutatóintézet, |
|
Állattani
és Állatökológiai Tanszék,
Gödöllő |
Méhtenyésztési
és Méhbiológiai Kutatócs., Gödöllő |
|
bakonyi.gabor
mkk.szie.hu |
bekesi
katki.hu |
|
|
|
|
Székács
András |
Papp
László |
|
az
MTA doktora |
az
MTA rendes tagja, kutatóprofesszor |
|
MTA
Növényvédelmi Kutatóintézete, |
Magyar
Természettudományi Múzeum, Budapest |
|
Ökotoxikológiai
és Környezetanalitikai O., Bp. |
lpapp
zoo.zoo.nhmus.hu |
|
aszek
nki.hu |
|
Írásunk
reakció Kiss József és munkatársai (2007)
környezeti kockázatelemző dolgozatára, amire
alapozva a vendégszerkesztő azt a következtetést
vonta le, hogy más rovarokat, mint a kukoricamolyt nem
veszélyeztet a vizsgált, géntechnológiai
úton módosított (GM) növény. Ezzel
szemben mi csak a GM-növények esetről esetre való
értékelésével érthetünk
egyet, hiszen nagyon is eltérnek az egyes fajtacsoportok
környezeti konzekvenciákkal járó
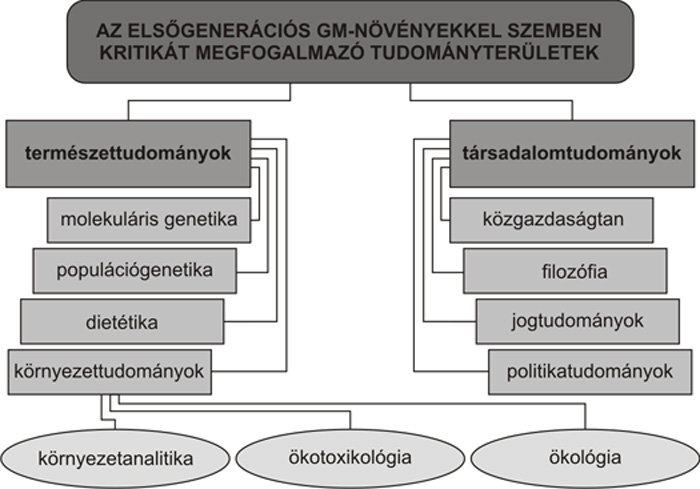
tulajdonságai. Korántsem csak a környezettudományok
kritizálják az első generációs
GM-növényeket, hanem más természet- és
a társadalomtudományok is (1.
ábra). A természettudományokon belül
a molekuláris és populációgenetika,
továbbá a dietétika fogalmazott meg eddig –
a környezettudományokon túl – megfontolandó
érveket.
Környezetanalitika
A
Cry-toxint termelő növények vizsgálatával
foglalkozva korántsem elhanyagolható, hogy a Bacillus
thuringiensis mely toxinját termeli egy fajta,
hiszen ennek rovarrendre kiterjedő hatása van. A toxinok közül
a gazdaságilag jelentősebbeket kiemelve az orális
hatású Cry1-toxinok a lepkefélék, a
Cry3-toxinok a bogarak, a Cry4-toxinok a kétszárnyúak
lárváit pusztítják elsősorban. Míg
a GM-növények e toxinok valamelyikét termelik, a
B. thuringiensis
törzsek 4–12 Cry-protoxint termelhetnek, amelyek a
gazdarovar emésztése során aktiválódnak,
majd kötődve a kevéssé ismert tulajdonságú
receptoraikhoz a bél hámsejthártyáját
roncsolják. E kapukon keresztül jut be a testüregbe
a bélben élő szimbionta mikrobaközösség,
ami szepszist idéz elő. Kiss és munkatársai
(2007) a Bt-kukorica
környezeti kockázatait értékelve e
tudományterületet egyáltalán nem érintik.
Cry1A-toxintermelő
képesség
– környezetanalitikai
vizsgálatok 1.
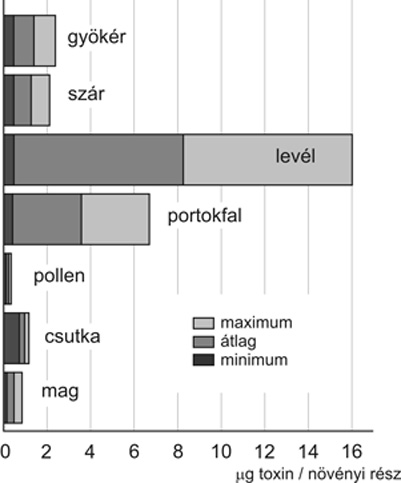
A
Bt-növények
által termelt Cry-toxinok mennyisége széles
határok között változik. Vannak genetikai
események, amelyekből származó növények
minden szövete azonos mennyiségű toxintermelésre
képes, s vannak, ahol szövetspecifikus toxintermelést
jegyeztek fel. A MON 810-es
fajtacsoportra, például egységnyi szövetre
vonatkoztatva levél > portokfal > gyökér >
szár > mag > pollen (2.
ábra); míg a megtermelt szerves anyagra
kalkulálva levél > szár > gyökér
> mag > portokfal > pollen sorrend írható fel.
Külön
problémaként említhető, hogy az azonos genetikai
eseményből előállított fajtákban a
Cry1A-toxintermelés jelentősen eltérhet, sőt ugyanazon
fajta – pl. növénytáplálás-függő
– különbségeket is mutathat. Az őszig
termelődő Cry1A-toxintartalom, a DK-440 BTY fajtában ~80 %-ban
a levelekben található (Székács –
Darvas in: Darvas,
2007). A DK-440 BTY által hektáronként
megtermelt Cry1Ab-toxintartalmat lehet az ötféle
Cry-toxint tartalmazó DIPEL rovarölőszerrel kiszórt
protoxin (80 % Cry1- és 20 % Cry2-toxin) mennyiségével
összevetni, ám a teljes mennyiségek (protoxin vs.
aktív toxin) összehasonlítását
megnehezíti, hogy előtte számtalan környezetanalitikai
alapkutatási problémát kell tisztázni. Az
egyik lényegi nehézség, hogy a kereskedelemben
kapható mennyiségi mérésre alkalmas ELISA
készletet a gyártója (EnviroLogix Inc.)
visszavonta a piacról, illetve ma már csak kvalitatív
mérésre forgalmazza.
Cry1A-toxin
lebomlása
a
tarlómaradványokban
– környezetanalitikai
vizsgálatok 2.
A
növényi sejtekbe zárt Cry1A-toxin aktivitása
a talajban legalább 234 napig megmarad (Tapp – Stotzky,
1998). A Cry1A-toxintartalom 1–8 %-a mérhető vissza egy
év múlva (Székács – Darvas In:
Darvas, 2007). Szárított növényi minták
hűtött körülmények között évekig
és szinte változatlan Cry1A-tartalommal tárolhatók.
Egy
orális hatású fehérje/lektin-toxin
minden, az érzékenyek körébe tartozó
rovart megbetegít, ami táplálkozással azt
magához veszi. Ezek a rovarok a kukoricát vagy
tarlómaradványait fogyasztják. Másodlagos
érintettek azok, amelyek kukoricán élő állatokat
fogyasztanak. Hatásnak kitett csoportok azok is, amelyek
tápláléka – a pollenszórás
időtartama alatt – szennyeződik a kukoricapollen által a
környezetben szétterülő Cry-toxinnal. A
környezetanalitikai vizsgálatok tehát jelzik azt,
hogy a MON 810-es
fajtákkal változó mennyiségű, főhatásként
a lepkehernyók megbetegítésére alkalmas
Cry1-toxin termelődik, majd tarlómaradvány formájában
hosszú ideig a területünkön marad.
Ökotoxikológia
Az
ökotoxikológiai vizsgálatok közül csak a
szabadföldről gyűjtött növényminták
ellenőrzött körülmények közötti,
gerincteleneket érintő laborvizsgálatára térünk
ki. Mindezt rizikóvizsgálatok előzik meg, amelynek
során az érzékenynek gondolt kapcsolatokat
kiválasztjuk. Megjegyzendő viszont, hogy a hatásvizsgálatok
környezetanalitikai háttér nélkül való
végzése vakrepüléshez hasonlítható,
hiszen a minták Cry-toxintartalmának nagymérvű
variabilitása miatt (2.
ábra) az oksági kapcsolatra egyébként
nem derülhet fény.
Kártevők
– főhatásvizsgálat. A MON
810 által termelt Cry1Ab-toxin hatását
legeredményesebben a levélre tojást rakó,
kelés után azon táplálkozást kezdő
kukoricamoly hernyóra fejti ki. A fejlődő szemeken károsító
gyapottok-bagolylepke hernyóin a hatás csekélyebb
(Kiss et al., 2007), amelynek oka szerintünk az, hogy ott egy
nagyságrenddel kevesebb Cry1A-toxin fejeződik ki (2.
ábra). A kukorica egyéb hazai kártevőire
a Cry1A-toxin hatásspektruma nem terjed ki. Így a
kukoricabogár, a pattanóbogár, a kukoricabarkó,
a fritlégy, a muharbolha, a levéltetű és a
kétfoltos takácsatka kártétele
zavartalan. Az Európa más tájain károsító
déli kukoricabagoly magyarországi kártételéről
viszont nincs tudomásunk. Kukoricamoly ellen egyébként
a hazai gazdák nem védekeznek, mivel a fertőzöttség
mértéke igen ritkán éri el az ún.
kártételi küszöbszintet.
Néhány
elemzés felveti annak lehetőségét, hogy a
kukoricamoly és a gyapottok-bagolylepke hernyóinak
csőkártétele a csőfuzariózis kialakulásához
teremti meg a lehetőséget. Ezt vizsgálatokkal kellene
tisztázni.
Védett
állatok – mellékhatás-vizsgálat 1.
A Cry1-toxin hatásspektruma a lepkefélék
hernyóira terjed ki. Magyarország védett
állatfajai között 187 lepkefajt találunk. A
DK-440 BTY pollennel termelt Cry1A-toxinmennyiség a táblaszéli
gyomokon élő kilenc védett lepkefajunkat (Darvas et
al., 2004) érintheti. Tehát nem csupán azt a
kettő, modellállatként választottat, amit Kiss
és munkatársai (2007) nekünk tulajdonítva
említ. A hazai kukoricatábla-szegélyeken a
harmadik leggyakoribb növény – növénykapcsolat
a nagy csalán, melyen két védett (nappali
pávaszem és atalantalepke) és egy ritka faj
(c-betűs lepke) hernyói kelhetnek akkor, amikor a hazai
kukoricafajták a pollenjüket szórják. A
tábla szegélyén a nappali pávaszem
hernyók esetében ~20 %-os pusztulás is
előfordulhat, ami jelzés arra, hogy élőhelyük a
MON 810 pollenjétől
megváltozik. A természetvédelmi törvények
ezt tilalmazzák (Darvas et al., 2006), ezért olyan
rizikóanalízist végezni, amely megengedett
pusztulást vesz figyelembe: értelmetlen. A
modellválasztás azért kívánatos,
mert az összes érintett faj vizsgálata képtelenség
volna. Téves Kiss és munkatársainak (2007) azon
megállapítása is, hogy a laboratóriumi
eredmények túlértékelik a veszélyt,
mert ennek ellenkezője az, ami előfordul (Lang et al., 2007). A
laboratóriumban a rovarok számára optimális
környezetet biztosítunk (nincs hőmérséklet-ingadozás,
eső, UV-sugárzás stb.), és a sűrű hálók
távol tartják a nagyobb testű ragadozókat és
a parazitoidokat. A patogének is figyelmet érdemelnek,
amelyek bármilyen időszaki legyengülés miatt
aktiválódhatnak. A cypovírus-2 együtt él
a nappali pávaszem hazai állományával, s
járványos formában söpörhet végig
a népességein. Védett lepkefajokon kapott
eredményeinket Lauber Éva és munkatársai
(Darvas, 2007) összegzik.
Hasznos
állatok – mellékhatás-vizsgálat 2.
E területről bizonyosan kiemelhetők a megporzást
végző méhek. A kukorica ugyan nem mézel, de
pollenjét a méhek kilométerekről begyűjtik,
felhasználják a fiasítás táplálására,
ezenkívül bekerül a mézbe is. A nyári
virágmézben megjelenő GM-kukoricapollen nemcsak a
bioméhészetek problémája, de az
exportorientált magyar méztermelésé is.
Néhány eddigi vizsgálat azt mutatta, hogy maga a
Cry1A-toxin közvetlenül nem mérgező a méhekre,
de megváltoztathatja viselkedésüket (Babendreier
et al., 2005). A DK-440 BTY pollen csökkenti a lárvák
fejlődési erélyét, illetve a kifejlett méhek
rezisztenciájára (Nosema
apis egysejtű parazita fertőzés) negatív
hatással van (Békési In: Darvas, 2007). Az egyéb
hasznos állatokkal (ragadozók és parazitoidok)
laboratóriumban végzett vizsgálatok száma
igen alacsony, és az egyes vizsgálatok átlagosan
nem haladták meg a három hét időtartamot (Lövei
– Arpaia, 2005).
Rezisztencia
– mellékhatás-vizsgálat 3. A
szubletális dózisban adott vegyületekre
szelektálódó rezisztens népesség
megjelenése általános jelenség az
élővilágban. Így van a DIPEL esetében is.
Vizsgálatainkban a tizedik nemzedékre szelektálódott
Cry1-toxinra (DK-440 BTY levélre) rezisztens
aszalványmoly-népesség. Ez azt veti fel, hogy az
egyféle toxint termelő Bt-növényekre
alapozó növényvédelmi megoldások
gyors lejáratúak (Darvas – Lauber In: Darvas,
2007). Ennek kezelésére még nem látunk
jól hasznosítható megoldásokat.
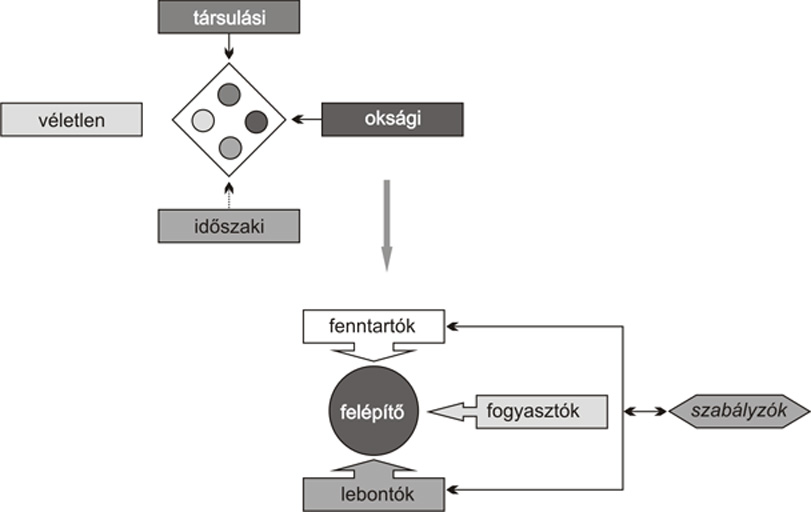
Ökológia
E
meglehetősen összetett egység vázlatos leírására
vállalkozunk csupán. Az ökológiai hatások
alatt szabadföldieket értünk, amelyeket az érintett
életközösség bonyolult rendszerén
próbálunk mérni. A bemutatáshoz Szelényi
Gusztáv praktikusan egyszerű csoportosítását
használjuk alapként (3.
ábra).
Egy
növényt jellemez, hogy élőhelyét időben
változóan más növényekkel osztja
meg. Kultúrnövényeknél leggyakoribb
szomszédja is a fajtársa. Azonban itt sem csak ilyen
kapcsolatok alakulnak ki. Jellemzően mások a táblaszéli,
illetve a táblán belüli kultúrnövény
nélküli foltok kapcsolatai. Pollenszóráskori
felvételezéseink során a hazai kukoricások
jellemző gyomjainak a táblaszegélyen a közönséges
tarackbúzát > a csillagpázsitot > a nagy
csalánt és > a vadkendert találtuk. Ugyanezen
állományok a vadköles > zöld muhar >
csattanó maszlag és > csillagpázsit
borítottsági sorrendet mutatták (Darvas et al.,
2004).
Amennyiben
egy növény állatkapcsolatait vizsgáljuk,
úgy oksági, társulási (együtt élő
növényekkel való természetes szabályzó
és tarlómaradvány-lebontó
kapcsolatmegosztás), időszaki (az év pár hetére
korlátozódó forrásra épülő
időszaki kapcsolat, például pollen- és
nektárfogyasztók) és véletlen (útban
lévő, megpihenő egyedek) kapcsolatokat találunk. Az
oksági összeköttetés alaptípusai: a
növényt fenntartó (megporzó), fogyasztó
(kártevő), lebontó (tarlómaradványi),
valamint az előzőeket szabályzó (ragadozók és
parazitoidok) kapcsolatok.
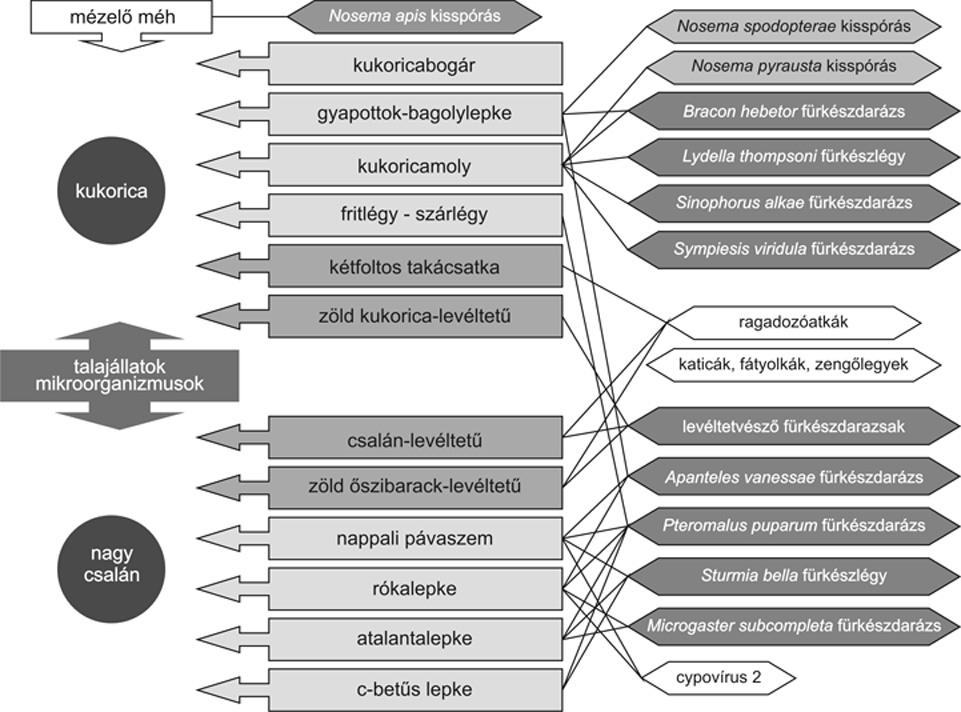
Ezek
az egyszerű táplálékláncok aztán a
növénytársulásokban bonyolult
táplálékhálózatokká
fonódnak össze. A kukorica – nagy csalán
kapcsolatra a mellékelt vázlat készíthető
el (4. ábra).
Természetesen ez messze bonyolultabb lesz, ha
valamennyi kapcsolatot feltüntetjük, illetve számolunk
a nagyobb élettérben megjelenő gerinces
táplálékláncokkal, melyek egyedi
lakóhelyén, ürülékén és
tetemén speciális közösségek alakulnak
ki.
Növény–növény
kapcsolatok
– mellékhatás-vizsgálat
4.
E
körbe sorolhatjuk a faj- és fajtahibridek keletkezésének
esélyét, amire egy koegzisztenciatörvénynek
támaszkodni kell. Úgy gondoljuk, hogy amíg
Bálint Andor (1980) állításait
kísérletesen nem cáfolja valaki, addig
vetőmagtermesztésnél 800 méterre tehető a
biztonságos izolációs távolság.
Minden ennél kisebb távolság tudományosan
megalapozatlan. Az izolációs távolságok
megválasztásának szigorúságával
ugyanakkor csupán egy gén terjedésének
gyorsaságát befolyásolhatjuk. Kukoricánál
800 méteres izolációs távolság
betartása sem biztosíték egy gén
terjedésének kiküszöbölésére,
csupán ésszerű kompromisszum, azaz osztjuk a világ
ökológusainak azon állítását,
hogy egymásra hatás nélkül maradó
növényi koegzisztencia – különösen
idegenbeporzóknál –: nonszensz.
Növényállományi
növény–állat kapcsolatok
– mellékhatás-vizsgálat
5.
Kiss
és munkatársai (2007) eredményei ebbe a körbe
esnek. Mielőtt erre rátérnénk, visszaidézzük
az évtizedekkel ezelőtti, az MTA NKI szervezésében
indult kukoricaökoszisztéma-vizsgálatok
eredményeit. Az 1976–1980 között hetenként
mintavételező – fény-, talaj-, szín-,
ragadós-, szexferomon-csapdákra épülő,
hálózást és növényvizsgálatot
alkalmazó – faunisztikai munka hosszú fajlistát
eredményezett (Mészáros, 1984). A
kultúrsivatagnak gondolt kukoricából 582
fonálféreg- és ízeltlábúfajt
mutattak ki. További meglepetés akkor ér
bennünket, ha átnézzük ezt a listát,
amelynek jó, ha 1-2 %-a kukoricakártevő, vagy a
kukorica tarlómaradványát bontja, és
talán 2-3 %-a tartozik a szabályzók körébe.
Honnan jön a többség? Nos, úgy tűnik, az
sokkal inkább arra a helyi, mikroszinten sokféle,
generalista életközösségekre jellemző, ahová
a kukoricát vetették. További megjegyzéseket
is biztonságosan tehetünk, s ezek a felvételezés
módjára (talajbeli felvételezések és
kinevelések korlátozottsága), a felvételezések
időpontjára (későiek hiányoztak) vonatkoznak.
Számos, feltáratlan csoport világos jelzés
a velük foglalkozó specialisták hazai hiányára.
Kiss
és munkatársai (2007) az előbbi munka nyomdokain
elindulva, kisparcellán, különböző csapdák
imágófogásaiból igyekeztek többnyire
generalista ragadozókra következtetéseket levonni
arról, hogy a Bt-növényeknek
van-e valamilyen állatközösséget átalakító
hatásuk. Adataik nem jeleztek szignifikáns eltéréseket.
Azonban a fajdiverzitás-rendezési görbék
önmagukban még nem jelentenek eredményt, csak azt
bizonyítják, hogy jogosult a diverzitások
sokféle indexszel való összehasonlítása.
Bemutatott eredményeik nem feltétlenül azt
jelentik, hogy eltérések nem lehetnek, csupán
arra utalnak, hogy az alkalmazott módszerek ezek
megállapítására nem teremtették
meg a lehetőséget. A 30x30 méteres parcellák
(Szekeres et al., 2006) nem alkalmasak jelentős helyváltoztató
képességű rovarok népességingadozásainak
követésére. A jól röpülő
rovarokra jellemző az élettartam alatti átlagosan 3–10
kilométeres szétterjedési távolság.
A kabócák közölt adataiból
kiszámítható a fajazonossági index, amely
2002-re 42 %, míg 2003-ra vonatkozóan 52 %. Ezek
két-két hasonló kinézetű, hazai
ökoszisztéma fajazonossági index értékei
lehetnének. Az eredmény persze nem azt jelenti, hogy a
kicsi, egymásba fűzött parcellák kabócái
között ekkora különbség van, csupán
azt, hogy a begyűjtött anyag elégtelen volt elemzésre.
Hogy az átrepülő kabócák közül
hány olyat vonzott a sárga ragacslap, amelyeknek semmi
közük a kukoricához, nem tudhatjuk.
A
kísérleti beállítás számos
más ponton is koncepcionálisan téves. A
vizsgálatokban meghatározó a floémból
táplálkozó levéltetvekre épülő
ragadozók követése, azonban ha a növényevő
szervezetébe a sejtekben lokalizálódó
Cry-toxinból igen kevés jut be (Dutton et al., 2002;
Burgio et al., 2007), akkor ragadozóján sem várható
hatás. Az alkalmazott csapdák talajszinten mászó
(például futóbogarak) és jól
röpülő (például katicabogarak, fátyolkák,
poloskák stb.) imágókat fogtak; ezek
szervezetéből viszont nem mutattak ki speciális
receptorokat, amelyen keresztül a Cry1-toxinok a hatásukat
kifejthetnék. Nem véletlen, hogy a Cry3-toxint termelő
kukoricafajták csak a kukoricabogár lárváit
pusztítják, míg az imágók
károsodás nélkül táplálkoznak.
Kiss és munkatársai (2007) vizsgálataiban a
fogott állatok töredékének lehet oksági
köze a kukoricához. A vizsgált polifág
állatok érintettségének behatárolása
az egyes zsákmányállatok Cry1-toxintartalmának
és fogyasztási hányadának mérése
nélkül képtelenség. A vizsgálatban
felsorolt nem-célállatcsoportok (például
pókok) nem tartoznak a Cry1-toxin hatáskörébe,
azaz csupán indirekt (nem a főhatás-mechanizmusán
alapuló) hatások keresése lehet az efféle
vizsgálat célja. Indirekt hatások természetesen
előfordulhatnak, például a táplálkozási
preferencia területén, de ezek – jó
hatékonysággal – csak tenyészeteken
vizsgálhatók.
Közleményük
futóbogarakra vonatkozó részét Szekeres
és mtsai (2006) cikkére alapozták. Miután
azonban vizsgálataikban nem volt a kukoricán kívüli
kontrollmintavétel, senki sem tudhatja, hogy a bogárfajok
közül melyiknek van köze a kukoricához. Maguk
is elismerik (a citált cikk 274. oldala), hogy a fajgazdagság
lehetséges oka a szomszédos gyümölcsösökből
való bevándorlás.
Hiányolható
az eredménymegvitatásból az a nagyon egyszerű
ökológiai szemlélet is, hogy ha egy kártevőt
kiemelünk egy közösségből, azzal annak
természetes szabályozóit is érintjük.
Nem maradhat tehát hatás nélkül az az
eljárás, amely kiemeli a kukoricamolyt a kártevők
közül, hiszen annak specialista szabályzói,
itt például a kukoricamoly-fürkészlégy
népessége vele fog csökkeni.
Talajbeli
életközösségek
– tarlómaradvány
– állatkapcsolatok
– mellékhatás-vizsgálat
6.
A
kukoricából többféle úton juthat
Cry-toxin a talajba. Ezek közül kétségtelenül
a gyökérváladékokon keresztül és
a tarlómaradványokkal a betakarítás után
talajba kerülő toxinmennyiség a legjelentősebb. A toxin
lebomlása területenként jelentősen eltérő,
elsősorban a talajélettől, a hőmérséklettől és
a talajtípustól függően. A hagyományos és
a Cry-toxint termelő kukorica beltartalma több vegyületet
tekintve is szignifikánsan különbözhet.
Mindezekből következik, hogy a Cry-toxint termelő kukorica
talajállatokra gyakorolt hatása térben és
időben jelentősen különböző lehet, valamint direkt és
indirekt hatásokkal is számolnunk kell. Ezeket a
tényeket komoly súllyal kell figyelembe venni a
laboratóriumi vizsgálatok eredményeinek
extrapolálása során és a terepvizsgálatok
eredményeinek elemzésekor.
A
Cry-toxin esetében az a kérdés, hogy a kukorica
által termelt toxinnak van-e letális vagy szubletális
hatása a talajállatokra. A fajtatulajdonosok
nullhipotézise az, hogy ilyen hatás nincs, és
véleményük szerint erre vonatkozó
tudományos adatokat sem publikáltak még. Viszont
más kutatási eredmények arra utalnak, hogy ez az
állítás nem tartható. Adatok bizonyítják
(1. táblázat), hogy
a Cry-toxint termelő kukorica izogénes párjához
viszonyítva szignifikáns hatást gyakorolt
talajállatokra. Ne felejtsük: jelen kérdésfeltevés
esetén érdektelen, hogy sok más esetben valóban
nem találtak szignifikáns különbségeket.
Mivel
a vizsgálatok célszervezetei, módszerei,
kísérleti beállításai,
talajtípusai stb. rendkívül heterogének,
általánosításra, összefoglaló
következtetések levonására nem alkalmasak.
Azt azonban határozottan állíthatjuk, hogy
számos, megalapozott kutatási eredmény
bizonyítékai szerint hatások léteznek.
Talajbeli
életközösségek – tarlómaradvány
– mikrobiális
kapcsolatok
– mellékhatás-vizsgálatok
7.
Ez
a legismeretlenebb része a kapcsolatoknak, hiszen becslések
szerint jó, ha a talajban élő mikroszervezetek fajainak
1 %-át ismerjük. Talaj-mikrobiológiai ismereteink
lényegesen kisebbek annál, mint ami az egyik
legfontosabb megújuló erőforrásunk, a talaj
okszerű műveléséhez, egészségének
megőrzéséhez szükséges lenne. Évszázadok
óta úgy hatunk erre a közegre (kezeljük
például mutagén hatású növényvédő
szerekkel), hogy annak élő alkotóelemeit nem tudjuk
megnevezni.
Utószó
Azzal
a fölöttébb egyszerű állítással
zárjuk sorainkat, hogy véleményünk szerint
nem írható le a szakmai reputáció
megőrzésének igényével az a mondat, hogy
nem merült fel a GM-növényekkel szemben megalapozott
környezettudományi aggály, vagy hogy
valószínűsíthető az, hogy ilyen eredmény
nem is fog születni egy körültekintő, oksági
vizsgálatsorozatban. Az is állítható,
hogy a felsorolt tíz vizsgálattípus eltérő
kérdésekre ad választ, így ezek egymással
nem helyettesíthetők. Környezetanalitikai alapok nélkül
a közölt biológiai eredmények kérdőjelesek.
Bizonyosra vehetjük, hogy a Bt-növényeknek
van nem kívánt környezeti hatásuk, s csupán
a kiterjedés és mérték az, ami ökológusok
között vita tárgyát képezheti. A
fentiekből levonható módszertani következményeket
a GMO-Kerekasztal szakemberei vizsgálati tervben foglalták
össze ( http://www.vedegylet.hu/doc/AllasfoglalasH.pdf
). Véleményünk szerint ezek elvégzése
után jelenthető ki bármi is az egyes GM-fajták
környezetre gyakorolt hatásáról.
Kulcsszavak:
MON 810, Bt-kukorica,
GM-növények, környezetanalitika, ökotoxikológia,
ökológia
IRODALOM
Babendreier,
Dirk – Kalberer, M. N. – Romeis, J. – Fluri, P. –
Mulligam, E. – Bigler, F. (2005): Influence of Bt-transgenic
Pollen, Bt-toxin and Protease Inhibitor (SBTI) Ingestion on
Development of Hypopharingeal Gland in Honeybees. Apidologie. 36,
585–594.
Bakonyi
Gábor – Szira F. – Kiss I. – Villányi
I – Seres A. – Székács A. (2006):
Preference Tests with Collembolans on Isogenic and Bt-maize. European
Journal of Soil Biology. 42, 132–135.
Bálint
Andor (1980): A vetőmagtermesztés genetikai alapjai.
Mezőgazdasági, Budapest, 171.
Burgio,
Giovanni – Lanzonia, A. – Accinellia, G. –
Dinellia, G. – Bonettia, A. – Marottia, I. –
Ramillia, F. (2007): Evaluation of Bt-toxin Uptake by the Non-target
Herbivore, Myzus persicae (Hemiptera: Aphididae), Feeding on
Transgenic Oilseed Rape. Bulletin of Entomological Research. 97,
211–215.
Darvas
Béla (szerk.) (2007): Mezőgazdasági géntechnológia
– elsőgenerációs GM-növények. Magyar
Országgyűlés Mezőgazdasági Bizottsága,
Bp., 164
Darvas
Béla – Csóti A. – Gharib, A. –
Peregovits L. – Ronkay L. – Lauber, É. –
Polgár A. L. (2004): Adatok a Bt-kukoricapollen és
védett lepkefajok lárváinak magyarországi
rizikóanalíziséhez. Növényvédelem.
40, 441–449.
Darvas
Béla – Székács A. – Bakonyi G. –
Kiss I. – Biró B. – Villányi I. –
Ronkay L. – Peregovits L. – Lauber É. –
Polgár A. L. (2006): Az Európai Élelmiszerbiztonsági
Hivatal GMO Paneljének a magyarországi
környezetanalitikai és ökotoxikológiai
vizsgálatokkal kapcsolatos állásfoglalásáról.
Növényvédelem. 42, 313–325.
Dutton,
Anna – Klein, H. – Romeis, J. – Bigler, F. (2002):
Uptake of Bt-toxin by Herbivores Feeding on Transgenic Maize and
Consequences for the Predator Chrysoperla carnea. Ecological
Entomology. 27, 441–447.
Kiss
József – Szekeres D. – Tóth F. –
Szénási Á. – Kádár F.
(2007): Genetikailag módosított növények és
környezeti kockázatok: A „Bt-kukorica”
példája. Magyar Tudomány. 167, 428–436.
Lang,
Andreas – Lauber, É. – Darvas, B. (2007): Early
Tier Tests are not Sufficient for GMO Risk Assessment. Nature
Biotechnology. 25, 35–36.
Lövei
L. Gábor – Arpaia, S. (2005): The Impact of Transgenic
Plants on Natural Enemies: A Critical Review of Laboratory Studies.
Entomology Experiments and Applications. 114, 1–14.
Mészáros
Zoltán (szerk.) (1984): Results of Faunistical Studies in
Hungarian Maize Stands. Acta Phytopathologica Academiae Sci. Hung.
19, 65–90.
Szekeres
Dóra – Kádár F. – Kiss J. (2006):
Activity Density, Diversity and Seasonal Dynamics of Ground Beetles
(Coleoptera: Carabidae) in Bt- (MON 810) and Isogenic Maize Stands.
Entomologica Fennica. 11, 269–275.
Tapp,
Hazel – Stotzky, Guenther (1998): Persistence of the
Insecticidial Toxins from Bacillus thuringiensis subsp. kurstaki in
Soil. Soil Biology and Biochemistry. 30, 471–476.
ábra
• Az első
generációs
GM-növényekkel szemben kritikát megfogalmazó
tudományterületek.
2.
ábra • A MON 810-ben mért
Cry1-toxinmennyiségek (AGBIOS database; CFIA DD, 1997; Wraight
et al., 2000; ANZFA TR, 2001; EPA, 2001, Bruns – Abel, 2003;
EFSA, 2004; Chilcutt – Tabashnik, 2004; Abel – Adamczyk,
2004; Lang et al., 2004; Nguyen, 2004; EFSA, 20061). A
termőhelyek
és az évjáratok okozta különbségek,
ill. a mintázás időpontja
hozzájárulnak a nagymértékű
szóráshoz.
 3.
ábra • Egy növény (felépítő)
kapcsolatvázlata
3.
ábra • Egy növény (felépítő)
kapcsolatvázlata
 4.
ábra • A kukorica – nagy csalán társulás
oksági kapcsolatai. (Megjegyzés: halványszürke
négyszög – rágó kártevők;
sötétszürke négyszög – szívó
kártevők;
halványszürke hatszög – betegségek;
fehér hatszög – ragadozók; sötétszürke
hatszög – parazitoidok.)
4.
ábra • A kukorica – nagy csalán társulás
oksági kapcsolatai. (Megjegyzés: halványszürke
négyszög – rágó kártevők;
sötétszürke négyszög – szívó
kártevők;
halványszürke hatszög – betegségek;
fehér hatszög – ragadozók; sötétszürke
hatszög – parazitoidok.)
|
|
Célcsoport |
Mire
hatott a növény? |
Hatás
jellege |
Szerző |
|
Laboratóriumi
vizsgálatok |
|
|
|
|
|
|
Ászkarák |
korai
pusztulás, növekedés |
pozitív |
Escher
et al. (2000) |
|
|
Ászkarák |
táplálékfogyasztás |
negatív |
Wandeler
et al. (2002) |
|
|
Földigiliszta |
testtömeg |
negatív |
Zwahlen
et al. (2003) |
|
|
Földigiliszta |
kelési
siker |
negatív |
Vercesi
et al. (2006) |
|
|
Ugróvillás |
táplálékválasztás |
negatív |
Bakonyi
et al. (2006) |
|
|
Ikerszelvényes |
ürülékmennyiség |
pozitív |
Weber
– Nentwig (2006) |
|
Üvegházi
vizsgálat |
|
|
|
|
|
|
Egysejtűek
és fonálférgek |
denzitás |
pozitív |
Griffiths
et al. (2006) |
|
Terepvizsgálatok |
|
|
|
|
|
|
Egysejtűek
és fonálférgek |
denzitás |
negatív |
Griffiths
et al. (2005) |
|
|
Fonálférgek |
táplálkozási
csop. aránya |
megváltozott |
Manachini–Lozzia
(2002); |
|
|
|
|
|
Arndt
(2006) |
- 1.
táblázat • Cry-toxint termelő kukorica
szignifikáns hatása talajállatokra (Az
irodalomjegyzékben nem található hivatkozások
Bakonyi Gábortól kérhetők el, illetve
megtalálhatóak a cikk internetes változatának
végén a Magyar
Tudomány honlapján:
./07aug/13b.html
)
1
Az irodalomjegyzékben nem található
hivatkozások Lauber Évától kérhetők
el, illetve megtalálhatóak a cikk internetes
változatának végén a Magyar
Tudomány honlapján:
./07aug/13b.html