Gyepvegetáció akklimatizációja emelt légköri szén-dioxid-koncentrációhoz:
hosszú időtartamú kísérletek eredményei
Nagy Zoltán
kandidátus, PhD, habil. egyetemi docens,
Szent István Egyetem
Növénytani és Ökofiziológiai Intézet
Nagy . Zoltan ![]() mkk . szie . hu
mkk . szie . hu
Szente Kálmán
kandidátus
Magyar Vállalkozásfejlesztési KHT.
Tuba Zoltán
DSc., mb. intézetigazgató, egyetemi tanár
SZIE Növénytani és Ökofiziológiai Intézet
MTA-SZIE Növényökológiai Kutatócsoport
A jelenlegi 380 ppm körüli légköri CO2-koncentráció század végére várható jelentős növekedése még a CO2-kibocsátási nemzetközi egyezmények betartása esetén is várható. A globális klímaváltozás egyik antropogén okaként megjelölt, emelkedő CO2-koncentráció azonban nemcsak az üvegházhatás révén befolyásolja a klímát, hanem a szárazföldi vegetáció megváltozó működése miatt is (Tuba et al., 2003). Itt elsősorban a növényzet csökkent párologtatására kell gondolnunk, amit a magasabb CO2-koncentráció a növényi gázcserenyílások vezetőképességének (a sztóma nyitottsági fokának) csökkentése révén fejt ki. Ennek egyenes következménye, hogy a levelek felszíni hőmérséklete megemelkedik, illetve hogy a növényzettel fedett felszínek esetében az energiamérlegben a szenzibilis hőáramra jutó hányad nagyobb lesz. A probléma (a globális klímaváltozás és a növényzet kapcsolatát tekintve) összetettségére jellemző, hogy a vegetáció fentebb említett csökkent párologtatásának jelentős szerep jut abban, hogy a folyók tengerekbe történő vízszállítása az utóbbi évtizedekben erősen megnövekedett (Gedney et al., 2006). Magyarországon a nyári csapadék nagyrészt záporok és zivatarok formájában hullik le, amelyek kialakulásában a felemelkedő levegő vízgőztartalma fontos szerepet játszik. Ha a levegő vízgőztartalma – a vegetáció emelt szén-dioxid-szint miatti csökkentett, „leszabályozott” vezetőképessége és párologtatása miatt − csökken, akkor a záporok és zivatarok kialakulásának esélye is csökken (Horváth, 2005), ami a nyári aszályok, hőséghullámok előfordulásának valószínűségét növeli. A 2003-as év nyarának súlyos aszályát részben a téli csapadék elmaradása és az ennek következményeként csökkent felszíni párologtatás okozta (Nagy et al., 2007). Bár ez utóbbi esemény nem köthető közvetlenül az emelkedő légköri CO2-koncentrációhoz, a hatás – az aszály kialakulása – hasonló, függetlenül attól, hogy a vegetáció csökkentett párologtatását a téli csapadék hiánya, vagy a magasabb légköri CO2-szint okozza. A CO2 ugyanakkor a növények növekedésének „alapanyaga” is, és mint ilyen, számos, a növények anyagcseréjét, így termésmennyiségét és annak összetételét közvetlenül befolyásoló élettani folyamatra is hatással van (Tuba, 2005). A sztóma-vezetőképesség már említett csökkenése nem csak a párologtatásra, hanem a növények CO2-asszimilációjára is hatással van (Sage et al., 1989; Tuba et al., 1996; Nagy et al., 1997). Ez a hatás azonban kettős: egyrészt csökkentő (a gázcserenyílások kisebb vezetőképessége miatt), másrészt pedig pozitív, elsődlegesen a CO2-megkötés enzimjének, a rubsicónak a serkentése miatt. Ehhez kapcsolódik, hogy kisebb enzimmennyiség és/vagy aktivitás is elegendő a fotoszintézis sebességének fenntartásához, és a fotoszintetizáló szövetek nitrogénkoncentrációja – a csökkent rubiscotartalom következtében – emiatt sok esetben csökken. Mindez maga után vonja, hogy emelt CO2-koncentráció alatt a növekedés nitrogénigénye csökkenhet. A termesztett növények hozamára, produkciójára az emelt légköri CO2-szint általában pozitív hatással van (Rogers et al., 1983). A megtermelt szénhidrát-mennyiséget azonban fel is kell „használni”, például a raktározó szerepű növényi szervekben, ellenkező esetben ugyanis a megnövekedett oldható cukortartalom negatív visszacsatolással csökkenti a fotoszintézis sebességét (Azcon-Bieto, 1983). Emiatt a megnövekedett légköri CO2-szint elsődlegesen azokra a növényfajokra lehet pozitív hatással, amelyek a fotoszintézis megnövekedett sebességét a raktározó szervekbe történő transzporttal képesek hasznosítani. A levélfelület-indexre gyakorolt hatás sem egyértelmű, ugyanis nő a levelek vastagsága (a specifikus levéltömeg), a levelek felülete így hasonló vagy akár megnövekedett levéltömeg mellett is változatlan maradhat vagy csökkenhet. A növények egyedi válaszai az emelkedő légköri CO2-koncentrációra tehát összetettek, függenek az adott növényfaj alaktani és élettani sajátságaitól is. A válaszok megismerését és a predikciót nehezíti, hogy a fajok többsége rövidebb-hosszabb kezelési időtartam után akklimatizálódik a megemelt CO2-szinthez (Makino, 1994; Nagy et al., 1997), amely válasz részben a már fent említett fotoszintézis-leszabályozásból, részben alaktani, allometriai módosulásokból áll, de – a növényi állomány szintjén – megváltoznak a vegetáció elsődleges funkcióit – anyag- és energiaforgalmát – a környezeti tényezők (források) függvényében leíró kapcsolatok is. Többfajú növényállományokban az állomány hozamára gyakorolt hatás összességében (az állomány egészére nézve) semleges vagy negatív is lehet (Amthor, 1995). Ilyen többfajú növényállományok alkotják a legeltetéssel vagy kaszálással hasznosított gyepek többségét is.
A gyepvegetáció részaránya a terresztris vegetációban meghaladja az egyharmadot, ezért mind a klímavédelem, mind a hasznosítás szempontjából fontos, hogy ismerjük a gyepvegtáció-típusok emelkedő légköri CO2-koncentrációra adott, várható válaszait. A gyepek jelentősége a fás vegetációval borított területek zsugorodásával várhatóan emelkedik. Ennek ellenére, amíg az erdei ökoszisztémák C-tároló szerepéről és kapacitásáról tekintélyes mennyiségű információ áll rendelkezésre, a gyepek vonatkozásában ez nem mondható el. Egyáltalán nagyon kevés ismeretünk van a gyepek működéséről, globális C-körforgalomban betöltött szerepéről, azok hosszú időtartamú emelt CO2-koncentrációra adott válaszairól. Pedig a gyepökoszisztémák az erdőökoszisztémáktól lényeges eltérő tulajdonságokkal bírnak. Ilyen például az, hogy amíg az erdőkben a szén felhalmozódása főképp a föld feletti, addig a gyepekben az a föld alatti biomassza formájában történik.
A kutatás célja az emelkedő légköri CO2-koncentráció hosszú távú növényökológiai hatásainak megismerése egy jellegzetes Kárpát-medencei (homokpusztagyep) és egy kelet-európai gyepvegetáció (löszpusztagyep) példáján. A vizsgálatok a vegetáció szerkezetének, illetve a vegetáció és növények fiziológiájának, produkciójának és ökofiziológiájának a vizsgálatára terjedtek ki. A munka a GATE (SZIE) Növénytani és Ökofiziológiai Intézete korábban EU-pályázati pénzügyi forrásokból felépített és működtetett „Globális klímaváltozás és növényzet” EU long-term kísérletes ökológiai kutatóállomásán (EU-programok és GCTE tagja) biztosított feltételekkel folyt.
Az expozíciós technika
A növények emelt légköri CO2-szint alatti neveléséhez felül nyitott tetejű, műanyag falú, 1,28 m átmérőjű, 1 m magasságú kamrákat (OTC), illetve a CO2-koncentráció emelését kamra nélkül biztosító, 1,5 m átmérőjű mini FACE-gyűrűket (Free Air CO2 Enrichment – FACE) alkalmaztunk. Az OTC-rendszerben a kamra alján körben elhelyezkedő ventillált polietilén csőben emeljük meg a befújt levegő CO2-koncentrációját, ami a cső falába vágott nyílásokon keresztül jut a kamra terébe, majd a kamra nyitott tetején távozik. A mini FACE-rendszer esetében a működés ezzel analóg, a különbség a kamra hiánya és az ebből eredő sokkal nagyobb CO2-felhasználás, illetve fluktuáló CO2-koncentráció. A kamrahatás (a kamra lényegesen melegebb környezeténél és a légáramlás szárító hatása is jelentős) elkerülése végett mégis érdemes volt ezt a technikát alkalmazni.
Az OTC-rendszerben 1994-től folytattunk kísérleteket transzplantált lösz- és homokpusztagyep monolitokon. A mini FACE-rendszer felépítése (a CO2-ellátó rendszer, az emelt CO2-koncentrációjú gyűrűk kontroll-rendszere, elektromos vezetékek, mikrometeorológiai állomás telepítése és üzembe helyezése) és a löszgyep monolitok transzplantációja 1998 júniusában fejeződött be, a fumigáció 2000 végéig tartott. A mini FACE-rendszerben a CO2-szint mellett két N-műtrágyázási szint is a kísérlet része volt.
A vizsgált objektumok
1.) Salvio Festucetum rupicolae löszpusztagyep eredetihez közeli és degradált állománya és fontosabb fajai: Festuca rupicola, Filipendula vulgaris, Salvia nemorosa; 2.) Festucetum vaginatae danubiale homokpusztagyep-állomány és fontosabb fajai: Festuca vaginata, Silene otites, Koleria glauca, Thymus marschallianus, valamint zuzmó (Cladonia convoluta, C. furcata), moha (Tortula ruralis) telepei.
Eredmények
Levélszintű nettó CO2-felvétel, akklimatizáció. A levél CO2 gázcseréjének egyenlegét a bruttó fotoszintézis és a (mitokondriális és fény-) légzés különbségeként a nettó fotoszintézis adja meg. Mérése infravörös gázanalizátorral egybekapcsolt levélkamrában történik, értéke függ a környező levegő CO2-koncentrációjától is, annak emelkedő értékeivel növekszik, a kontroll- és az emelt CO2-koncentráción nevelt növényeknél egyaránt. A kezelésnek tulajdonítható különbség a görbék lefutásában keresendő, nevezetesen a kezdeti emelkedés és a maximum (a telítési érték) értékeiben. A maximumnak a kontrollhoz hasonlított nagyobb értéke esetén a növény – levélszinten – pozitívan válaszolt az emelt CO2-szintre, emellett a válasz lehet semleges vagy negatív is a fotoszintézis leszabályozása miatt. A pozitív válasz azt jelenti, hogy az adott faj valószínűen sikeresebb lehet az emelt CO2-koncentrációjú légkörben, míg a negatív válasz ennek ellenkezőjét sejteti. A válasz élettani háttere többtényezős, a leszabályozást (a fotoszintézis–CO2-koncentráció görbe telítési szakaszának gátlását) ugyanis okozhatja a relatív foszforhiány éppúgy, mint a megtermelt szénhidrátokat fogadni képes raktár hiánya. Rövid (a fajtól függően néhány hónapos, egy-két éves) expozíciók esetén általában a pozitív válasz volt jellemző a vizsgált esetek túlnyomó többségében, az emelt CO2-koncentráción nevelt növények fotoszintézis-[CO2] görbéje magasabbra futott, mint a kontrollnövények esetében. Hosszabb (öt éves) expozíció után azonban a válasz a fajok egy részénél lefelé módosult (leszabályozás), azaz a kezelt és a kontrollnövények hasonló lefutású görbét mutattak, más részüknél viszont a fenntartott pozitív akklimatizációs válasz volt jellemző. E két esetet tapasztaltuk homokpusztagyepi és löszpusztagyepi (1. ábra) fajoknál egyaránt, azaz a válasz független volt a gyep típusától, ellenben függött attól, hogy a vizsgált faj pázsitfűféle vagy pedig kétszikű faj volt. A fűfajok ugyanis a fotoszintézis leszabályozását, a kétszikűek pedig a hosszabb időtartamon át is fenntartott pozitív akklimatizációt mutatták. Miután a két talaj tápanyagellátottsága erősen különbözik, a válaszok hasonlósága a két gyep esetében inkább köthető a megemelt szénhidrátszint általi visszacsatolás jelentkezéséhez (fűfélék) vagy annak elmaradásához (kétszikűek), mint a P-hiányhoz. Azaz a kétszikű fajok esetében a raktárak (pl. a gyökérrendszer) nagyobb kapacitásuk révén hosszú expozíció után is alkalmasak voltak a többlet-szénhidráttartalom hasznosítására, míg a fűfajok esetében nem. Utóbbiaknál a legnagyobb raktár maga a növekedési folyamat lehet, gyökérrendszerük ugyanis kevésbé alkalmas raktározásra, mint az évelő kétszikűeké, maghozamuk tömegaránya (teljes földfeletti tömeghez képest) pedig kicsi. Ezek az eredmények az OTC-ben nevelt növényekre vonatkoznak, de hasonló eredményeket kaptunk a mini FACE-rendszerben tartott növények esetében is.
További, a fentiekkel összhangban lévő eredményt az oldható cukor- és szénhidráttartalom vizsgálata hozott (mini FACE, löszgyep), ahol azt tapasztaltuk, hogy a fűfaj (Dactylis glomerata) és a kétszikű faj (Filipendula vulgaris) ellentétes képet mutatott a hajnali szénhidrátszintek alakulásában (2. ábra). A hajnali mintavételt az indokolja, hogy intenzív éjszakai felhasználás (transzport/növekedés) esetén hajnalra az oldható cukrok és a keményítő mennyisége csökken, ellenben a növekedés (vagy általában a szénhidrátokat „fogyasztó” folyamatok) hiányában említett metabolitok szintje kevéssé változik. A legyezőfű (F. vulgaris) leveleiben az oldható cukrok szintje kisebb volt a kezelt állományban, mint a kontrollban, a napközbeni hasonló aktivitás (esti értékek) ellenére, azaz a felhasználás sebessége a kezelt növények esetében valószínűleg nagyobb volt. A fű esetében ellentétes képet kaptunk, a szénhidrátok éjszakai felhasználásának mértéke a kontrollnövényekben nagyobb volt, mint az emelt CO2-koncentráción tartottakban. A szén-dioxid-gázcserében és szénhidrátmintázatban mutatott képet a hajtások, illetve levelek növekedési sebessége is alátámasztotta, azaz a füvek növekedési sebessége csökkent, a kétszikűeké pedig növekedett az emelt CO2-szint hatására. A növekedési válasz egy további fontos információt ad, ha a gázcserében és a szénhidrátmintázatban tapasztaltakkal együtt vesszük figyelembe; ez pedig az, hogy a kétszikű fajok a levél-, illetve a hajtásnövekedés szintjén is kedvezőbben reagálnak, mint az egyszikűek, az összetett levélszerkezet plasztikusabb válasz lehetőségét biztosítja. Másként fogalmazva a hajtás növekedése (és nem csak a raktározó szerveké) maga is pótlólagos szénhidrát-felhasználást jelent, elkerülve ezzel az egyébként (a füvek esetében) a fotoszintézisre a szénhidrátszinten keresztül megvalósuló negatív visszacsatolást.
Az emelt CO2-koncentráció hatására általában csökken a levelek fehérje-nitrogén tartalma, illetve a klorofillok, karotinoidok (A-vitamin prekurzor) mennyisége is. Ugyanakkor, legalábbis a fotoszintézisükben leszabályozott fajokban nő a szénhidráttartalom, és többnyire fokozódik a rostképződés. Mindez pedig közvetlenül befolyásolhatja a növény–állat interakciót, az állatok táplálkozását, anyagcseréjét és viselkedését, valamint természetesen az emberi táplálkozást is.
Állományszintű válaszok
Az eddigiekben egyes növényfajok levélszintű válaszait vizsgáltuk, és az ezekből származó eredmények azt valószínűsítik, hogy az egyszikű fajok valószínűleg hátrányba kerülhetnek a kétszikűekkel szemben a mainál magasabb légköri CO2-koncentráció mellett. Ezek az eredmények, bár értékesek lehetnek, nem adnak választ arra a nagyon is kézenfekvő kérdésre, hogy a vegetáció egésze hogyan válaszolhat az emelt CO2-szintre. Ha erre a kérdésre nem is nyerhettünk választ, a vizsgált gyep növényállományának viselkedésére nézve az alábbi eredményeket kaptuk.
Az emelt CO2-koncentráció a kontrollkezeléshez képest a sokfajú gyep állományának szintjén vizsgálva:
• jó csapadékellátású évben növelte az összes (földterületegységre vonatkoztatott) biomassza mennyiségét,
• csökkentette a levélfelület-index értékét,
• a vegetáció felszíni hőmérsékletének 1,5-2 °C-os emelkedését okozta,
• a kétszikű fajok borításának növekedését eredményezte az egyszikűek hátrányára.
A termesztett növények esetében általánosan tapasztalthoz hasonlóan az emelt CO2-szint a sokfajú gyepvegetáció esetében is növelte az egységnyi földterületre eső biomassza mennyiségét, bár nem szignifikáns mértékben. Ehhez hozzátartozik, hogy ez csak egy viszonylagosan jó csapadékellátású évben volt igaz, szárazságstressz alatt (a 2000-es évben) már nem volt különbség a kezelések között.
A levélfelület-index az 1 m2-re eső levelek (egyik oldalának) összfelületét jelenti. Becslése a lombozat fényelnyelésének mérésével lehetséges. Az említett LAI-csökkenés (3. ábra) valószínűleg az emelt CO2-szint bevezetőben már említett specifikus levéltömegnövelő hatásának, illetve részben a fűfajok csökkent növekedési sebességének tudható be, amely hatásokat az állomány szintjén a kétszikűek nagyobb növekedése – tehát nagyobb egyedi levélfelülete – sem volt képes ellensúlyozni. A csökkent levélfelület-index – a szűkebben tartott gázcserenyílások mellett – már elégséges volt a vegetáció felszíni hőmérsékletének szignifikáns emelkedéséhez a kontrollállományhoz képest (4. ábra). Tehát a működési (sztómák, fotoszintézis és növekedés) és alaki (specifikus levéltömeg és állományszinten a levélfelület-index) jellemzők együttes változásai egy olyan választ – a növényállomány kisebb párologtatását és nagyobb felszíni hőmérsékletét – eredményeztek az állomány szintjén, amely már nagy térléptékben is fontos és részben felelős lehet olyan jelenségekért, mint például a folyók megemelkedett vízhozama. Utóbbi esetben a csökkentett párolgás miatt az elfolyás – lokálisan esetleg nem észlelhető mértékben – növekszik, ami egész vízgyűjtők területét figyelembe véve viszont már szignifikáns mennyiség lehet.
A kétszikű fajok borításának növekedése és az egyszikűek borításának párhuzamos csökkenése – az eddig tárgyaltakkal összhangban – olyan eredmény, amely közvetlenül mutatja, melyik csoport lehet előnyben a másikkal szemben a várható klímaváltozás során.
Kulcsszavak: globális klímaváltozás, növénytömeg, akklimatizáció, állományszintű válaszok, fotoszintézis, vízhasznosítási efficiencia
Irodalom
Amthor, Jeff. S. (1995): Terrestrial Higher-Plant Response to Increasing Atmospheric [CO2] In Relation to the Global Carbon-Cycle. Global Change Biology. 1, 243–274.
Azcon-Bieto, Joaquin. (1983): Inhibition of Photosynthesis by Carbohydrates in Wheat Leaves. Plant Physiology. 73, 681–686.
Gedney, Nicola – Cox, P. M. – Betts, R. A. – Boucher, O. et al. (2006): Detection of a Direct Carbon Dioxide Effect in Continental River Runoff Records. Nature. 439, 835–838.
Horváth Ákos (2005): A gomolyfelhőktől a hurrikánokig – a konvekció mint a légkör egyik legfőbb bizonytalansági tényezője. Magyar Tudomány. 7, 797–804.
Makino, Amane (1994): Biochemistry Of C3-Photosynthesis in High CO2. Journal of Plant Research. 107, 79–84.
Nagy Zoltán – Pintér K, – Czóbel Sz. et al. (2007) The Carbon Budget of a Semiarid Grassland in a Wet and a Dry Year in Hungary. Agriculture Ecosystems and Environment. 121, 21–29.
Nagy Zoltán – Szente K. – Tuba Z. (1997): Acclimation of Dicot and Monocot Temperate Species to Long-Term Elevated CO2 Concentration. Abstracta Botanica. 21, 329–336.
Rogers, Hugo H. – Thomas, J. F. – Bingham, G. E. (1983): Response of Agronomic and Forest Species to Elevated Atmospheric Carbon Dioxide. Science. 220, 428–429.
Sage, Rowan F. – Sharkey, T. D. – Seeman, J. R. (1989): Acclimation of Photosynthesis to Elevated CO2 in Five C3 Species. Plant Physiology. 89, 590–596.
Tuba Zoltán (ed.) (2005): Ecological Responses and Adaptations of Crops to Rising Atmospheric Carbon Dioxide. Haworth Press Inc, New York, USA
Tuba Zoltán – Szente K. – Nagy Z. – Koch J. (1996): Responses of CO2 Assimilation, Transpiration and Water Use Efficiency to Long-Term Elevated CO2 in Perennial C3 Xeric Loess Steppe Species. Journal of Plant Physiology. 148, 356–361.
Tuba Zoltán – Raschi, A. – Lannini, G.M. et al. (2003): Vegetations with Various Environmental Constraints under Elevated Atmospheric CO2 Concentrations. In: Sanità di Toppi, Luigi – Pawlik-Skowronska, Barbara (eds.): Abiotic Stresses in Plants. Dordrecht, Kluwer Academic Publishers, 15–204
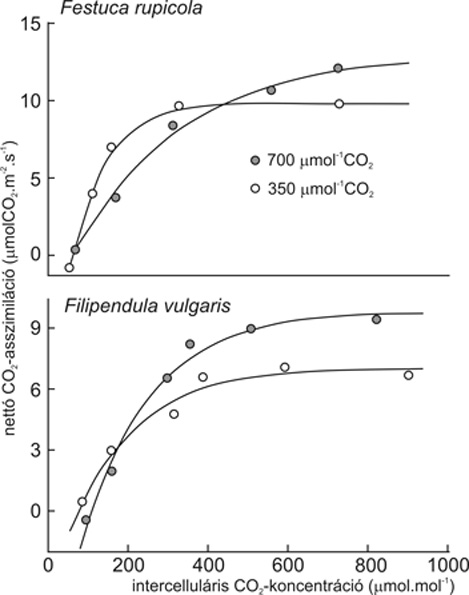
1. ábra • A nettó fotoszintézis CO2-koncentráció-függése két löszgyepi faj, a Festuca rupicola és a Filipendula vulgaris 350, illetve 700 µmol.mol-1 CO2-koncentráció mellett élő egyedeinek leveleiben ötéves expozíció (OTC) után
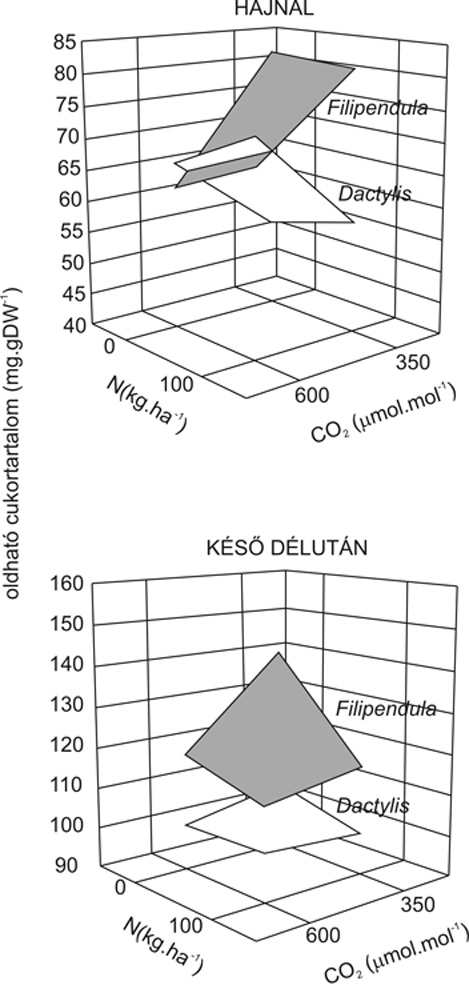
2. ábra • Az oldható cukrok hajnali és késő délutáni szintjei A Dactylis glomerata és a Filipendula vulgaris eltérő CO2-koncentráción (és N-ellátottság mellett) fejlődött egyedeinek levelében kétéves expozíció (mini FACE) után. A növekedési, illetve a transzportfolyamatok révén hajnalra a levelek cukortartalma csökken, így az említett folyamatok sebességeinek kezelések közötti kvalitatív összevetésére nyílik lehetőség. Késő délutánra az oldható cukrok szintjei megemelkednek, értékeik a hajnalban mérteknél magasabbak
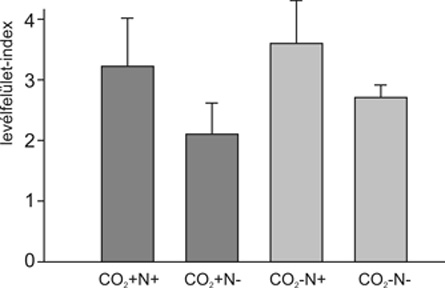
3. ábra • A levélfelület-index (LAI) értékei eltérő CO2-koncentráción (expozíció: mini FACE-rendszerben) és N-ellátottság mellett a löszgyep állományaiban az intenzív növekedési periódusban (április vége). Nitrogén hozzáadása növelte, a CO2-kezelés, viszont (1. és a 3., illetve a 2. és a 4. oszlopok összevetése) csökkentette a levélfelület-indexet
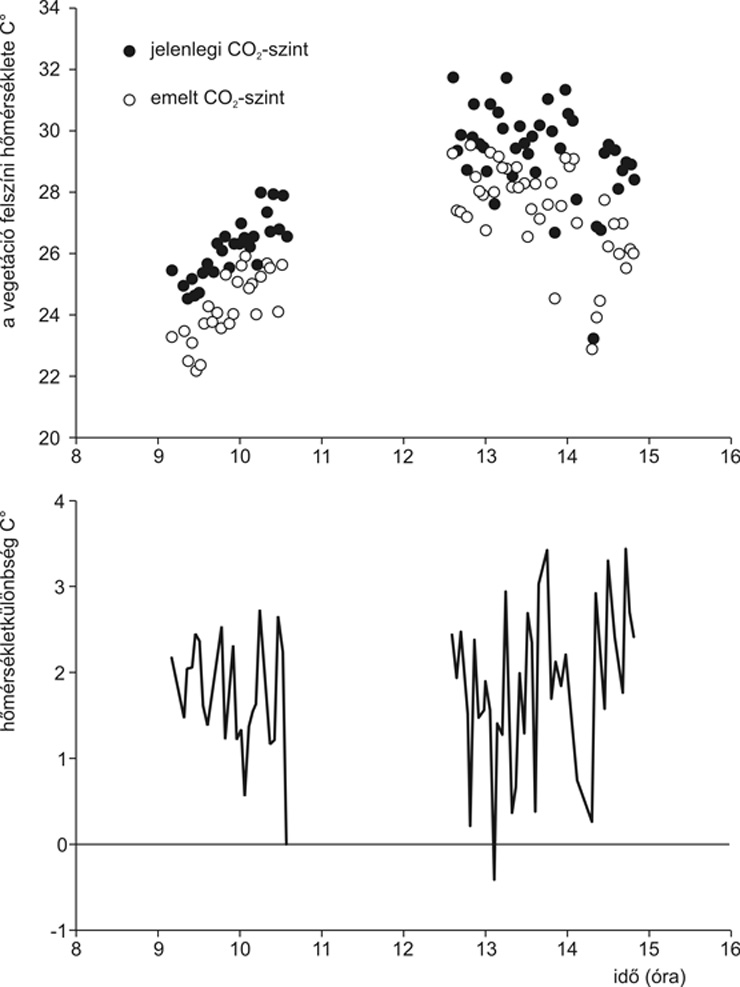
4. ábra • A vegetáció felszíni hőmérsékletének értékei a löszgyep eltérő CO2-koncentráción fejlődött állományaiban augusztus hónapban. Az egyes mérési pontok tíz mérés átlagai. Az emelt légköri CO2-koncentráció a sztómák szűkítése és az így csökkentett párologtatás révén a vegetáció felszíni hőmérsékletét csökkenti
<-- Vissza a 2007/10 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]