Mérsékelt övi gyepközösségek CO2-gázcseréjének
kisléptékű térbeli variabilitása és mintázata
Fóti Szilvia
PhD-hallgató, tanszéki mérnök
SZIE Növénytani és Ökofiziológiai Intézet
Foti . Szilvia ![]() mkk . szie . hu
mkk . szie . hu
Balogh János
PhD-hallgató, tudományos segédmunkatárs
MTA–SZIE Növényökológiai Kutatócsoport
Nagy Zoltán
kandidátus, PhD, habil. egyetemi docens
SZIE Növénytani és Ökofiziológiai Intézet
Czóbel Szilárd
PhD-hallgató, egyetemi adjunktus
SZIE Növénytani és Ökofiziológiai Intézet
Bartha Sándor
kandidátus, PhD, habil. tudományos tanácsadó
MTA Ökológiai és Botanikai Kutató Intézet
Tuba Zoltán
DSc, mb. intézetigazgató, egyetemi tanár
SZIE Növénytani és Ökofiziológiai Intézet
MTA–SZIE Növényökológiai Kutatócsoport
A vegetáció szerkezetét és annak megváltozását térben és időben különböző léptékekben vizsgálhatjuk. A gyepek kutatásának térbeli tartománya hagyományosan a kis lépték, mely néhány centimétertől néhány méterig terjed, mert ebben a tartományban nyilvánul meg a florisztikai kompozíció legnagyobb változatossága (Bartha et al., 1997).
A fátlan növényállományok, s ezen belül a nagy változatossággal bíró gyepek, így a mérsékelt övi gyepek fiziológiai tulajdonságai is léptékfüggően, a cönológiai kontextus alakulásának megfelelően változnak (Tuba et al., 1998).
A fiziológiai működések közül kiemelten fontos a gyepek fotoszintetikus teljesítménye és légzése, melyek ráadásul összefüggnek a gyepek globális szénkörforgalomban betöltött szerepével is, ugyanis a szárazföld felszínének kb. 40 %-át fátlan növényközösségek borítják. A CO2-gázcsere térben és időben nagy variabilitást mutat minden ökoszisztémában (Stoyan et al., 2000); ez a folyton változó környezeti feltételekhez való alkalmazkodás kulcsa.
Számos módszer létezik fátlan növényállományok CO2-gázcseréjének mérésére. A kisléptékű térbeli variabilitás nyomon követésére elsősorban különböző méretű és alakú, nyílt vagy zárt rendszerű gázcseremérő kamrák használatosak (Angell et al., 2001).
A variabilitás legegyszerűbben a mért változó variációs koefficiensével (CV) fejezhető ki. Két további módszer a térbeli mintázat vizsgálatára is alkalmas, s teljesen új az ökofiziológiában. Az egyik esetben növekvő mintavételi egységgel dolgozunk (egymásba ágyazott kvadrátok módszere), a másik esetben konstans méretű mintavételi egységeket növekvő szeparációs távolság mellett hasonlítunk össze (geostatisztikai módszer). Az előbbi módszer a CO2-gázcsere mérése esetében több, már említett, különböző méretű gázcseremérő kamra (1. ábra) (Czóbel et al., 2005) párhuzamos alkalmazását jelenti. E vizsgálatokhoz hét kamrából álló sort fejlesztettünk ki, melyek átmérője 7,5 cm-től 480 cm-ig duplázódik, s melyek mindegyike zárt, átlátszó és henger alakú. A geostatisztikai módszer esetében a fentiek közül egy kamra használatáról van szó, amellyel hetvenöt ponton, kijelölt kör (transzszekt) mentén mérünk. A minta mérése mindkét esetben hordozható infravörös gázanalizátorral történik.
A mért értékek csak akkor alkalmasak térbeli mintázatanalízisre, ha a kapott minta eleget tesz az állandósági (stacionaritási) feltételnek, valamint kellően nagy a minta mérete. A kamrasorozat alkalmazásával olyan karakterisztikus mintavételi egységnagyságot (karakterisztikus áreát) kereshetünk, melynél a gyep CO2-gázcseréje a legnagyobb vagy a legkisebb variabilitást mutatja. A legnagyobb variabilitást mutató egységet (maximumarea) használhatjuk a későbbiekben a mintázatanalízishez szükséges adatok összegyűjtéséhez, mert ezzel a kamramérettel feltehetőleg a CO2-gázcserének is nagy változatossága fogható meg, s így juthatunk a legtöbb ismerethez a gyep működéséről. A legkisebb variabilitással jellemezhető mintavételi egység a működés minimumareája.
A geostatisztika számos módszere közül az ökofiziológiai célokhoz a félvarianciát (szemivarianciát), illetve az ennek alapján álló félvariogramot (szemivariogramot) találtuk alkalmasnak. A mért értékek páronkénti eltérésnégyzeteinek kiszámításával, s növekvő szeparációs távolság függvényében történő ábrázolásával kapott telítési görbék szerint a közelebbi minták hasonlóbbak egymáshoz, mint a távolabbiak, majd egy bizonyos távolságtól függetlenné válnak egymástól. A szemivariogramok paraméterei alapján meghatározható a vizsgált változó karakterisztikus foltmérete, és becslés is végezhető az ismeretlen, meg nem mért pontokra modellezéssel (krigeléssel). A krigelt mintázatok allokálásával ábrázolható a vizsgált változók foltmintázata.
Munkánk néhány eredményt villant fel az állomány-fotoszintézis, a talajlégzés és néhány háttérben húzódó talajparaméter mikroskálájú térbeli variabilitásáról és mintázatáról, valamint az állományok működésbeli (szünfiziológiai) karakterisztikus léptékeinek meglétéről. Számos mérsékelt övi gyepökoszisztémán folytattunk méréseket, itt most közülük három különböző és földrajzilag is távol eső ökoszisztémáról, a bugaci extenzív legelőről, az isaszegi löszsztyeppről és a domb- és hegyvidéki mátrai gyepről (1. táblázat) származó eredményeket mutatunk be. Valamennyi helyszínen vizuálisan homogén, közel vízszintes helyzetű állományfoltokban mértünk, többnyire a gyepek maximális biomasszájú, kora nyári időszakában. E kutatásaink célja a funkcionális términtázatok első megismerése, egy mikroléptékű tér- és időbeli fotoszintézis-modell megalapozása, melyekkel sérülékeny, fragmentált hazai és más gyepökoszisztémák szénkörforgalomban betöltött szerepének jobb megismerését reméljük.
A mátrai extenzív és intenzív kezelésű gyepekben végzett szünfiziológiai mérések eredményei szép példát adtak arra, hogy kevéssé limitált időpontban, homogén gyepfoltban meghatározhatók az állomány karakterisztikus foltméretei. Az extenzív legelőn az összes mintavételi egységnagyságnál alacsonyabb nettó ökoszisztéma-gázcsereértékeket (NEE) mértünk, mint az intenzív kaszálón (2. ábra). A nettó fotoszintézis variabilitása fordított volt, nagyobb CV-t az extenzív gyep esetében kaptunk csaknem minden mintavételi egységnél, ami e terület nagyobb kompozíciós változatossága, vizuálisan is feltűnő jelentősebb foltossága alapján feltételezhető volt. A variabilitás minimuma extrapolációval volt becsülhető: a szünfiziológiai minimumarea: 625 cm-nek, illetve 1066 cm-nek adódott az extenzív és az intenzív gyepekben (10 % CV-re extrapolálva). E gyepekben a CV maximuma (maximumarea) a 30 cm átmérőjű mintavételi egységnél található.
A löszpusztagyepen folytatott vizsgálatok meglepő eredményt hoztak a CV léptékfüggése és a CO2-gázcsere allokált términtázata szempontjából. A nettó fotoszintézis (3. ábra) variációs koefficiense a 60 cm átmérőjű kamránál minimumot mutatott, ami feltehetően a társulás fotoszintézisének lokális minimum áreája. A geostatisztikai mintázatelemzéssel arra kerestük a választ, hogy mi a CV emelkedésének oka a 60 cm átmérőjű mintavételi egységnél nagyobb kamrák alkalmazásakor.
A geostatisztikai analízis során azt tapasztaltuk, hogy az állomány CO2-gázcseréjének a vizsgált térbeli tartományban nem volt észlelhető, robusztus, kis foltokból álló mintázata, amit amúgy mikrocönológiai struktúrája alapján vártunk volna. A talajlégzés és a talajvíztartalom allokált értékei és krigelt mintázatai alapján egyértelműen láthatóvá vált egy, a választott térbeli léptéken túlmutató foltosság jelenléte. A terület cserjésedése valószínűleg komoly befolyást gyakorolt az állományszintű fiziológiai működésre, hiszen a cserjék nemcsak közvetlen környezetükben, hanem akár jó pár méter távolságra is kihatnak a mikroklíma és számos talajtulajdonság alakulására (Breshears et al., 1997). Ez a jelenség magyarázhatta a több mintavételi egységnagysággal folytatott vizsgálatban a lokális, s nem abszolút variabilitásminimum jelentkezését is.
A geostatisztikai analízishez a bugaci homoki legelőn is a 30 cm átmérőjű kamrával végeztük a fotoszintézis- és légzésméréseket. A homoki gyepek mozaikos kompozíciójáról és mikrocönológiai szerkezetéről meglévő előzetes ismereteink alapján alig feltételeztük, hogy szép, statisztikailag szignifikáns paraméterekkel alátámasztott funkcionális términtázatot találunk majd, mégis ez történt.
A homoki legelő nettó fotoszintézis-, talajlégzés- és talajvíztartalom-mintázatai a választott térbeli felbontás léptékébe eső foltokkal voltak jellemezhetők (4. ábra), s e mintázatok szignifikáns szemivariogram paraméterekkel rendelkeztek. A „forró pontok” (maximumok) nagyjából ugyanarra a helyre estek. A homoki legelő tehát igen jól szervezett (szabályozott) és ezért kiváló objektumnak bizonyult a CO2-gázcsere térbeli mintázatának vizsgálatára.
Összefoglalás
Vizsgálataink valamennyi objektumon azt bizonyították, hogy a CO2-gázcsere variabilitása térben nem állandó, függ a mintavételi egység nagyságától és a mintavétel helyétől is.
Az átlagos CO2-gázcsere és annak variációs koefficienssel kifejezett variabilitása mellett lényeges új ismeretekhez, valós térbe allokált funkcionális mintázatokhoz jutottunk az alkalmazott módszerekkel. A CO2-gázcsere bemutatott foltmintázatai, karakterisztikus areái ugyanakkor csak „pillanatfelvételek” e gyepek fenológiai optimum-időszakából. A megismert mintázatok megváltozhatnak, évszakonként eltérő jellegzetességekkel rendelkezhetnek, megnyilvánulhat a vízhiány vagy hőstressz csakúgy, mint a degradáció vagy más dinamikai folyamat, ahogy erre eredményeink is utalnak.
Az itt leírt, a társulásszintű ökofiziológiai funkciók mintázataira és szerveződésükre vonatkozó vizsgálataink exploratív jellegűek. Fontos kiemelni, hogy ilyen típusú és ilyen részletességű terepi ökofiziológiai vizsgálatokat előttünk még sehol sem végeztek. Ugyanakkor várható, hogy a bioszféra globális változásai során legelőször éppen ebben, az általunk kiválasztott, néhány dm-től néhány m-ig terjedő állományszintű léptéktartományban lépnek majd fel kritikus – később más léptékekre is továbbgyűrűző – változások. A karakterisztikus foltok, mint akár „működési egységek” nagyobb, táji léptékű folyamatok alapjai, így a foltméretek megváltozása nagyobb terület ökofiziológiai funkcióit is érinti.
Kutatásainkat az OTKA-32586 project, továbbá a GREENGRASS, a CARBOMONT, a CARBOEUROPE és a MERCI EU-projektek támogatták.
Kulcsszavak: CO2-gázcsere, karakterisztikus area, krigelés, szemivariancia, térbeli mintázat, variációs koefficiens
Irodalom
Angell Raymond F. – Svejcar T. – Bates J. et al. (2001): Bowen Ratio and Closed Chamber Carbon Dioxide Flux Measurements over Sagebrush Steppe Vegetation. Agricultural and Forest Meteorology. 108, 153–161.
Bartha Sándor – Czárán T. – Scheuring, I. (1997): Spatio-temporal Scales of Non-equilibrium Community Dynamics: A Methodological Challange. New Zealand Journal of Ecology. 21, 2, 199–206.
Breshears, David D. - Rich, P. M. - Barnes, F. J. - Campbell, K. (1997): Overstory-imposed Heterogeneity in Solar Radiation and Soil Moisture in a Semiarid Woodland. Ecological Applications. 7, 4, 1201–1215.
Czóbel Szilárd – Fóti Sz. – Balogh J. et al. (2005): Chamber Series and Space-Scale Analysis in Grassland Vegetation. A Novel Approach. Photosynthetica. 43, 267–272.
Stoyan, Helmut - De-Polli, H. - Böhm, S. et al. (2000): Spatial Heterogeneity of Soil Respiration and Related Properties at the Plant Scale. Plant and Soil. 222, 1–2., 203–214.
Tuba Zoltán – Csintalan Zs. – Nagy Z. et al. (1998): Szünfiziológia: alapozó gondolatok és exploratív vizsgálatok egy születő növényökológiai tudományterülethez. In: Fekete Gábor (szerk.): A közösségi ökológia frontvonalai. Scientia, Budapest, 171–196.

1. ábra • Növekvő mintavételi egységek: 7,5 cm-től 480 cm-ig duplázódik a gázcseremérő kamrák átmérője
|
|
Bugac |
Isaszeg |
Mátra |
|
|
|
|
|
|
Évi átlagos csapadék (mm) |
500 |
550–600 |
600 |
|
Évi átlaghőmérséklet (°C) |
10,5 |
11 |
10,5 |
|
Tengerszint feletti magasság (m) |
140 |
230 |
300 |
|
Talaj |
csernozjom típusú homok |
humusz- és tápanyaggazdag homokos lösz |
köves, kötött agyag |
|
Domináns fajok |
Festuca pseudovina |
Festuca rupicola, Chamaecytisus austriacus |
Arrhenatherum elatius (kaszáló), Festuca pseudovina (legelő) |
|
Kezelés |
extenzív legeltetés húsz éve folyamatosan |
felhagyott, cserjésedik |
intenzíven kezelt kaszáló és extenzív művelésű legelő |
táblázat • A három vizsgálati helyszín néhány jellemzője
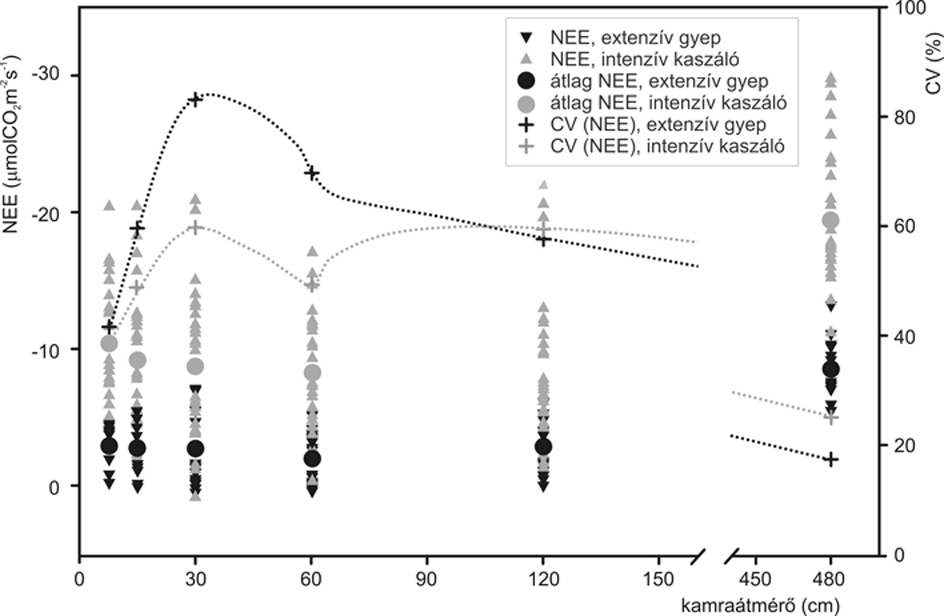
2. ábra • A nettó fotoszintézis átlagának (µmol CO2m-2s-1) és variációs koefficiensének (%) függése a mintavételi egység nagyságától a mátrai gyepekben
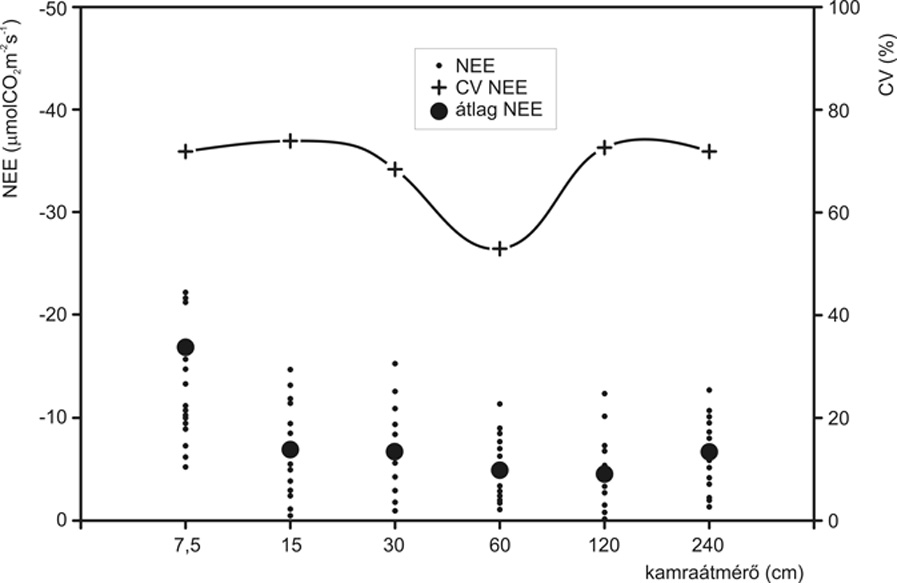
3. ábra • A nettó fotoszintézis átlagának (µmol CO2m-2s-1) és variációs koefficiensének (%) függése a mintavételi egység nagyságától löszpusztagyep esetében
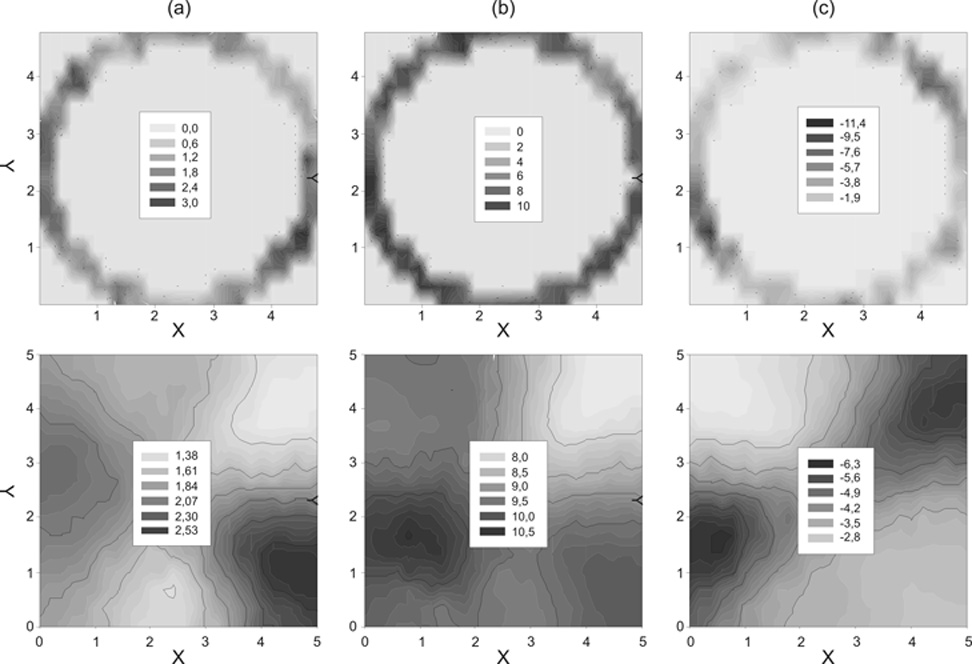
4. ábra • A talajlégzés (a, µmol CO2m-2s-1), a talajvíztartalom (b, %) és a nettó fotoszintézis (c, µmol CO2m-2s-1) mért (felső ábrák) és becsült (alsó ábrák) értékeinek allokált mintázata a bugaci homoki legelő esetében. A kör mentén megmért foltok talajlégzése és víztartalma a kisebb értékektől a nagyobbakig, a világosabbtól a sötétebb színekig terjedő színskálával ábrázolt foltmintázatot mutatta. A megmért értékeket becsléssel terjesztettük ki egy-egy nagyobb, 5×5 m-es foltra, majd ugyanúgy a szürketónusú színskálát rendeltük az értékekhez, mint a fenti két ábrán. Így vált láthatóvá egy nagyobb terület mintázata, mely mindkét tulajdonságra fokozatos átmenetet mutatott az egyik irányban, két eltérő típusú, nagyobb folt találkozására utalva
<-- Vissza a 2007/10 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]