Növényökofiziológia
Megváltoztatott
kezelésű
hazai
gyeptársulásaink
funkcionális
ökológiai válaszai
|
Czóbel
Szilárd |
Szirmai
Orsolya |
|
PhD-hallgató,
egyetemi adjunktus |
PhD-hallgató,tudományos segédmunkatárs |
|
SZIE
Növénytani és Ökológiai Intézet |
MTA–SZIE Növényökológiai Kutatócsoport |
|
czobel . szilard  mkk . szie . hu mkk . szie . hu |
|
|
|
|
|
Szerdahelyi
Tibor |
Nagy
János |
|
PhD,
egyetemi adjunktus |
PhD,
egyetemi adjunktus |
|
SZIE
Növénytani és Ökológiai Intézet |
SZIE
Növénytani és Ökológiai Intézet |
|
|
|
|
Balogh
János |
Fóti
Szilvia |
|
PhD-hallgató, tudományos segédmunkatárs |
PhD-hallgató,
tanszéki mérnök |
|
MTA–SZIE Növényökológiai Kutatócsoport |
SZIE
Növénytani és Ökológiai Intézet |
|
|
|
|
Péli
Evelin |
Pintér
Krisztina |
|
PhD-hallgató, tudományos segédmunkatárs |
PhD-hallgató,
tanszéki mérnök |
|
MTA–SZIE Növényökológiai Kutatócsoport |
SZIE
Növénytani és Ökológiai Intézet |
|
|
|
|
Horváth
László |
Nagy
Zoltán |
|
DSc,
habil. vezető főtanácsos |
kandidátus,
PhD, habil. egyetemi docens |
|
Országos
Meteorológiai Szolgálat |
SZIE
Növénytani és Ökológiai Intézet |
|
|
|
|
Tuba
Zoltán |
|
|
DSc,
mb. intézetigazgató, egyetemi tanár |
|
|
SZIE
Növénytani és Ökofiziológiai
Intézet |
|
|
MTA–SZIE
Növényökológiai Kutatócsoport |
|
Bolygónk
szárazföldjeinek mintegy negyedén gyepökoszisztéma
a potenciális természetes vegetáció,
melynek kb. 20 %-át – az Antarktisz kivételével
minden kontinensen előforduló – mérsékelt
övi gyepek alkotják (IUCN, 1999). A trópusi és
szubtrópusi övben a hőmérsékleti
viszonyok lehetővé teszik a vegetáció folyamatos
fejlődését, de a mérsékelt övben a
téli fagypont alatti vagy közeli hőmérséklet
nyugalmi állapotba (dormancia) kényszeríti a
növényeket. A kontinentális éghajlaton
kialakult gyeptársulásoknál, a nyári
aszályos időszakban megfigyelhető egy másik nyugalmi
periódus is, melynek oka nem a magas hőmérséklet,
hanem a vízhiány következtében fellépő
szárazságstressz (Archibold, 1995). Ebben az időszakban
a kevés csapadék és magas hőmérséklet
okozta sztómazáródás hatására
a napi szénmérleg a tavaszinál alacsonyabb
értéket mutat. A növények áttelelő
szervek segítségével élik túl a
kedvezőtlen időszakokat. Nyári vízhiány idején
a domináns, évelő füvek párhuzamosan
növelik gyökérzetük és csökkentik
hajtásuk növekedését, majd nyugalmi
állapotba kerülnek, összehajló levelekkel. A
hagymás és gumós fajok a kellően csapadékos
időszakokban virágoznak tömegesen, míg a
száraz és hideg periódust föld alatti
szerveik segítségével vészelik át.
Csapadékos években – a sivatagokhoz hasonlóan
– gyakoriak a magállapotban áttelelő, rövid
életciklusú efemer növények. A többnyire
fajgazdag füves élőhelyek niche szegregációjára
nemcsak a különböző funkcionális csoportok
(például életformák és
fotoszintézis-típusok) együttes jelenléte,
hanem eltérő felépítésű gyökérzetük
is utal, ami csökkenti a nedvességért vívott
kompetíciót.
A
mérsékelt övi füves ökoszisztémák
föld feletti biomasszájában jellemzően az összes
szerves szén kevesebb, mint 1 %-a található
(Burke et al., 1997).
Ennél lényegesen nagyobb a gyökérzetben
tárolt organikus szén mennyisége, de legnagyobb
széntároló kapacitással a talaj
rendelkezik, mely a füves ökoszisztéma összes
szervesszén-tartalmának döntő tömegét
(átlag 90 %) tárolja. Fentiekből következően
csak a légkör–vegetáció–talaj
rendszer összes komponensét vizsgáló,
integrált megközelítés képes
leírni megfelelően a gyepökoszisztémák
szénkörforgását. Napjainkig kevés
ilyen jellegű, komplex publikáció jelent meg, de a
rendelkezésre álló adatok azt igazolják,
hogy a vegetáció és a talaj széntartalma
jelentősen eltérhet különböző gyeptípusok
esetén (Reeder – Schuman, 2002). Ez elsősorban
klimatikus eltérésekre vezethető vissza, de edafikus
tényezők is okozhatják. Eddigi ismereteink birtokában
megállapítható, hogy az adott gyeptípus
szerves széntartalma az évi csapadékmennyiség
növekedésével pozitívan, míg az évi
középhőmérséklet emelkedésével
negatívan korrelál (például Burke et al.,
1989).
Az
eltérő fiziognómiájú, struktúrájú
és diverzitású hazai gyeptársulások
(Tuba et al., 2004b)
kiterjedése a korábban művelt területek rovására
az elmúlt néhány évtizedben jelentősen
megnőtt, szocioökonómiai okok miatt. Hazánk
csatlakozva az Európai Unióhoz vállalta 1 millió
hektár mezőgazdasági művelés alatt álló
terület hosszú távú felhagyását,
ezért még a közeljövőben is növekedni
fog a felhagyott területek aránya. EU-csatlakozásunknak
a hazai agrárgazdálkodásra gyakorolt
további várható hatása, hogy egyes
területeken (kaszálók, legelők) megváltozhat
a gyepek tradicionális kezelési intenzitása, így
a korábban extenzíven kezelt gyepek egy részén
intenzívebb művelés (például műtrágyázás,
öntözés) várható.
Az
említett változások nemcsak a gyepek cönológiai
viszonyaira és diverzitására hatnak, hanem
párhuzamosan megváltoztathatják a fajok
működését (Szente et
al., 1996) és az adott rendszer C- és
N-körforgását, ezáltal befolyásolva
a főbb üvegházhatású gázok fluxusait
is (Soussana et al., 2007). A füves területek
gázcseréjében szerepet játszó
három üvegházhatású gáz közül
a CO2-nál a talaj és a vegetáció
szerepe döntő, a N2O-ot a talajok, míg a
CH4-t a legelő állatok bocsátják ki,
de utóbbi a talajjal is cserélődhet (Soussana et
al., 2007). Az elmúlt évek hazai (Tuba et
al., 2004a; Balogh et
al., 2007) és nemzetközi (Soussana et
al., 2007) ökofiziológiai és
mikrometeorológiai kutatásai során
bebizonyosodott, hogy a vizsgált hazai gyepökoszisztémák
– hasonlóan a vizsgált európai füves
területekhez és leszámítva az extrém
száraz éveket – a talajjal és légkörrel
alkotott rendszerben mint nyelők játszanak szerepet az
üvegházhatású nyomgázok cseréjében.
Nagy kiterjedésű gyepterületeink tehát jelentős
szerepet töltenek be az üvegházhatású
gázok (továbbiakban GHG) Magyarországra vetített
éves mérlegében, hiszen döntően a
közlekedés és az ipar által kibocsátott
GHG egy szignifikáns részét megkötik.
Ezért is fontos, hogy minél több hazai
gyeptársulás globális szén- és
nitrogén-körforgalomban betöltött szerepét
megismerjük, valamint különböző manipulációs
kísérletekkel felkészüljünk az előre
jelzett földhasználati és klimatikus változások
funkcionális ökológiai hatásainak
predikciójára.
Kutatásunk
során két eltérő alapkőzethez köthető,
különböző jellegű (például fajkészlet,
fiziognómia, struktúra) hazai gyeptársulásban
vizsgáltuk három éven keresztül (2002–2004)
a legeltetés felhagyásának, a korábban
kezeletlen gyep műtrágyázásának, illetve
öntözésének az állományok
szénkörforgására és növényökológiai
viszonyaira gyakorolt hatásait. Jelen publikációban
bemutatjuk a vizsgált gyepökoszisztémák
diverzitásában, cönológiai és
fiziognómiai szerkezetében, főbb funkcionális
csoportjaiban, állományszintű működésében
(infravörös gázanalizátorral/CIRAS-2, PP
Systems, Hitchin, UK és ún. kamrás
technikával mért CO2-fluxus adatok alapján),
produkciójában (mennyiségi, minőségi),
továbbá a talaj szén- és
nitrogéntartalmában bekövetkező változásokat.
A több tudományterületet felölelő (például
meteorológia, botanika, ökofiziológia, talajtan)
sokrétű mérések és analízisek
a fluxusvizsgálatokkal párhuzamosan történtek.
A
kiválasztott objektumok közül a homokterületek
legeltetésével kialakult homoki száraz
legelő (Cynodonti-Festucetum
pseudovinae) Bugacpusztához közel, a
Kiskunsági Nemzeti Park területén található
(é. sz. 46o41’, k. h. 19o36’, tszf. magasság
113 m). A hazánk homokterületein jellegzetes másodlagos
növénytársulás enyhén lúgos
(pH 7,3–8,6) homoktalaja csernozjom típusú.
A kiválasztott sík területen egy kb. 6 ha-os
elkerített rész reprezentálta a legeltetés
felhagyását. A szürke marhával
extenzíven legeltetett területen az átlagos
legelési ráta 0,53 és 0,75 állat/ha
között változott.
A
homoki legelő mellett kiválasztott magas füvű,
széleslevelű kétszikűekben gazdag, diverz
löszgyeptársulás (Salvio-Festucetum
rupicolae) az eurázsiai sztyeppzóna
hazai képviselője. Termékeny talaja miatt hajdani
állományainak többsége intenzíven
művelt mezőgazdasági területté alakult. In situ
vizsgálatainkat a Gödöllői-dombság nagyobb
kiterjedésű löszgyepekkel jellemezhető területén
végeztük, melynek gyengén lúgos (pH
7,6–7,9) talaja közepesen erodált, mészlepedékes
csernozjom. Az Isaszeg és Nagytarcsa közötti
sztyeppréten (é. sz. 47o42’, k. h. 19o24’,
tszf. magasság 255 m) két transzszekt mentén
kiválasztott tíz állományfolt (1 m ×
1 m) közül ötben évi egyszeri alkalommal –
a vegetációs időszak kezdetén –
műtrágyáztuk a gyepet (a kiszórt mennyiség
100 kg N ha-1, 50 kg P ha-1, 50 kg K ha-1-nak felelt meg). Ebből a
löszgyepállományból a Szent István
Egyetem Botanikus Kertjébe (Gödöllő, é. sz.
47o36’, k. h. 19o26’, tszf. magasság 220 m)
transzplantált tíz monoliton végeztük
az öntözés hatását vizsgáló
kísérletet, öt-öt öntözött és
kontroll állományfolton (0,8 m × 0,8 m). A
vegetációs időszak alatt üzemelő, automata
öntözőrendszer – éjszakai locsolással –
biztosította a gyep előre meghatározott, egyenletes
vízellátottságát (0,35 m3/m3
talajvíztartalom alatt tartva az öntözött
gyepet).
A
manipulációs kísérletek eredményét
a vizsgálati időszak klimatikus anomáliái
befolyásolták. A bugaci homoki legelőn 2003 első
intenzív növekedési periódusában
(március–június) a lehullott csapadék
összmennyisége mindössze 40 %-a volt az ötvenéves
átlagnak. Ezzel szemben 2004 hasonló időszakában
a gyepre 3,5-ször több csapadék hullott, mint
2003-ban. Hasonló eltéréseket tapasztaltunk a
vizsgált löszgyepek csapadékviszonyainak
elemzésekor is, mely jelentős mértékben
befolyásolta a gyepek működését és
produkcióját. Az Európa-szerte extrém
száraz 2003-as esztendő (Ciais et al., 2005) a gyepek
fiziológiai aktivitása mellett befolyásolta a
bugaci területre jellemző tradicionális kezelést
(féléves folyamatos, extenzív legeltetés),
a legeltetést ugyanis 2003 nyarán két hónapra
(július, augusztus) meg kellett szakítani.
A
kiskunsági homoki legelőn a legeltetés felhagyása
kismértékben növelte a pillangósok relatív
arányát, de nagyobb mértékben
csökkentette az egyéb kétszikűekét. A
vegetációs periódusban havonként nyílt
kamrás módszerrel mért nettó ökoszisztéma
CO2-fluxus értékeink azt mutatják,
hogy egyes időszakokban a legelt részen nagyobb volt a
CO2-elnyelés, mint az elkerített területen.
Mindez különösen akkor említésre méltó,
ha figyelembe vesszük a legeltetés felhagyása
miatt bekövetkezett eltéréseket a
levélfelület-értékek között. Az
azonos mikrometeorológiai viszonyok között mért
CO2-fluxus, illetve az ebből számított
szénmérleg nem tért el szignifikánsan a
legeltetett és a legeltetéstől elzárt gyep
között a kutatási időszakban. Ennek feltehető
oka, hogy a legeltetéstől elzárt állomány
magasabb biomasszamennyiségét és zöld
levélfelületét kompenzálta a jelentősebb
avarfelhalmozódás, valamint a lebomlás
következtében fellépő magasabb légzésintenzitás,
továbbá a tápanyagok lassabb reciklizációja
és az elmaradt trágyázás.
A
homoki gyep CO2-megkötése az optimális
vízellátottságú és fenológiai
fázisú évben a vizsgált kontroll
löszgyepekhez hasonló volt (1.
ábra). A föld alatti biomassza tömege a
legeltetéstől elkerített állományhoz
képest átlag másfélszeresére
növekedett, ezáltal a legelt részen szignifikánsan
kisebb értéket kaptunk (1/20 és 1/25) a föld
feletti és föld alatti biomassza arányára.
A talaj szénakkumulációja a legeltetéstől
elzárt gyepnél mindhárom vizsgált szint
esetében kisebb mértékben nőtt, mint a legelt
területen, és ez hosszú távon
befolyásolhatja a fiziológiai működést.
A
löszgyep esetében mindkét kezelés
lecsökkentette a C4-es fajok borítását és
az életformák számát. (A döntően
trópusi, szubtrópusi területekről szétterjedő
C4-es növények többsége a 45º-nál
alacsonyabb füves élőhelyeken jellemző.) A vizsgált
funkcionális csoportok közül a műtrágyázás
megnövelte a kétszikűek relatív arányát,
míg öntözés hatására az
egyszikűek váltak monodominánssá, és
a pillangós fajok teljesen eltűntek a gyepből. A műtrágyázást
toleráló fajok a tápanyag-utánpótlás
hatására növelték levélfelületüket,
magasabbra nőttek, kompenzálva az eltűnt, illetve lassan
adaptálódó fajok hiányzó
levélfelületét és biomasszatömegét.
Az öntözés pozitív hatásaként
szárazságstresszelt időszakban közel négyszeresére
nőtt a zöld levélfelület a kontrollállományhoz
képest, emiatt szignifikánsan nagyobb (másfélszeres)
borítási értékeket mértünk az
öntözött vegetációnál. A
megfelelő vízellátottság következtében
az egyébként forró és csapadékhiányos
nyári hónapokban is jelentős CO2-megkötést
mértünk az öntözött gyepben. Az egyenletes
vízellátottság a C-fixáció
növekedéséhez képest jóval kisebb
mértékben növelte a talajlégzést.
Mindkét kezelt löszgyep CO2-felvételében
negatív visszacsatolás figyelhető meg, melynek
feltehető oka a gyors biomassza növekedés és
N-akkumuláció következtében a vegetációs
időszak előrehaladtával fellépő N-hiány volt. Az
öntözött növényzetnél a föld
feletti/föld alatti biomassza aránya szárazságlimitált
időszakban nagyobb volt a kontrollállományhoz képest.
Ennek oka a már említett intenzív föld
feletti biomasszanövekedés, mellyel – N-pótlás
hiányában – nem volt képes lépést
tartani a gyökérzet növekedése. A
csapadékhasznosítási rátánál
(éves föld feletti nettó primer produkció/évi
csapadékösszeg; részletesebben lásd Huxman
et al., 2004) megfigyelhető, hogy még az optimális
vízellátottságú öntözött
gyep működését is kismértékben, de
negatívan befolyásolta az aszály. Ennek
feltehető oka, hogy a gyepet döntően C3-as fajok uralják,
melyek sztómazáródása (a déltáji
alacsony RH miatt) csökkentette a produkciót. Csapadékos
évben az intenzív növekedés és az
ezt kísérő talaj–növényzet
allokáció miatt az öntözött gyep föld
alatti biomasszájának N-tartalma jelentősebben, míg
a talaj N-tartalma közel azonos mértékben csökkent
a kontroll- és az öntözött állománynál.
A biomassza C-tartalma, a földhasználati módtól
és kezeléstől függetlenül csak szűk
tartományban változott.
Vizsgálataink
alapján megállapítható, hogy az extrém
száraz periódus a szünbotanikai viszonyokat
és fiziológiai aktivitást tekintve egyaránt
nagyobb mértékben stresszelte a kevésbé
strukturált, de humuszban gazdag homoki gyepet, mint a
löszgyepet (1. ábra).
Szén-dioxid-gáz szempontjából
a kutatott löszgyep éves szinten – kezeléstől
függetlenül – megkötőnek tekinthető, míg
a kevésbé strukturált homoki gyep csak
extrém száraz években tekinthető CO2-forrásnak.
A vizsgált paraméterek közül a leggyorsabb
változás a fajkompozíció esetében
figyelhető meg, míg a CO2-gázcsere
intenzitása csak az öntözött és
műtrágyázott löszgyepnél tért el a
kontrollhoz képest, az aktuális talajvízviszonyok
függvényében. A diverzitás és a
cönológiai értékek fluktuációja
jelzi, hogy a szünbotanikai működés nagyobb
környezeti perturbáció (például
2003. évi aszály) után is helyreáll. A
talajparaméterek jellemzően csak hosszabb időskálán
változnak szignifikánsan, de trendek már három
év alatt is megfigyelhetők. A löszgyepeknél
kezelés hatására észlelt csökkenő
fajszám és diverzitás, elsősorban az erős
kompetitor fajok növekvő dominanciája (Salvia
nemorosa a műtrágyázott, Dactylis
glomerata az öntözött gyepben) miatt
következett be. A fajkompozíció és a
borítási értékek megváltozásából
nemcsak az egyes fajok érzékenységére
következtethetünk, hanem a jövőben sikeres taxonok
predikciója is lehetővé válik a manipuláció
során kiválasztott gradiens mentén. Kutatásunk
igazolta, hogy a kezelési eredmények értékelésénél
– és ezek modellezési alapadatként történő
felhasználásánál – figyelembe kell
venni egy kezdeti akklimatizációs periódust,
melynek hossza a kezelés jellegétől, a klimatikus
viszonyoktól és a vegetációdinamikai
folyamatok intenzitásától függ. Az
adaptáció során megfigyelhető gyors
vegetációdinamikai folyamatok nemcsak a cönológiai
viszonyokat változtatják meg, hanem az állomány
struktúráját, diverzitását és
LAI-értékeit is jelentősen módosíthatják.
A klimatikus eltérések lehetővé tették a
biomasszanövekedés és a csapadékmennyiség
közötti összefüggés vizsgálatát.
Megállapítottuk, hogy a gyeptípusok és
kezelések közül a csapadékhasznosítási
ráta csak a műtrágyázott gyepnél
növekedett lineárisan, míg a többi
gyeptípusnál telítődésjellegű volt.
Eredményeink rövid távon nem támasztják
alá azt a hipotézist, hogy a szünbotanikai
jellemzők megváltozásával a szünfiziológiai
működés is párhuzamosan változik.
A
szerzők ezúton köszönik meg a GREENGRASS (EU
Framework 5 project), CARBOMONT (EU Framework 5 project),
CarboEurope-IP (EU 6 Framework project), NitroEurope- IP (EU 6
Framework project), NKFP6-00079/2005 projektek, valamint a Scientia
Amabilis Alapítvány anyagi támogatását,
továbbá a Kiskunsági Nemzeti Park
együttműködését.
Kulcsszavak:
földhasználat,
homokpusztagyep, löszpusztagyep, diverzitás, felhagyás,
öntözés, műtrágyázás,
biomassza, szénmérleg
IRODALOM
Soussana,
Jean Francois – Allard, V. – Pilegaard K, et al. (2007):
Full Accounting of the Greenhouse Gas (CO2, N2O, CH4) Budget of Nine
European Grassland Sites. Agriculture, Ecosystems and Environment.
121, 1–2., 121–134.
Archibold,
O. William (1995): Ecology of World Vegetation. Chapman and Hall,
London,
Balogh
János – Nagy Z. – Fóti Sz. – Pintér
K. – Czóbel Sz. – Péli E. R. –
Acosta, M. – Marek M. V. – Csintalan Zs. – Tuba Z.
(2007): Comparison of CO2 and H2O Fluxes over Grassland Vegetations
Measured by the Eddy-Covariance Technique and by Open System Chamber.
Photosynthetica. 45, 2, 288–292.
Burke,
Ingrid C. – Laurenroth, W. K. – Milchunas, D. G. (1997):
Biogeochemistry of Managed Grasslands in Central North America. In:
Paul, Eldor A. – Paustian, K. – Elliott, E. T –
Cole, C.-V. (eds.): Soil Organic Matter in Temperate Agroecosystems:
Long-term Experiments in North America. CRC Press, Boca Raton,
85–102.
Burke,
Ingrid C. – Yonker, C. M. – Parton, W. J. et al. (1989):
Texture, Climate, Cultivation Effects on Soil Organic Matter Content
in US Grassland Soils. Soil Science Society of America Journal. 53,
800–805.
Ciais,
Philippe – Reichstein, M. – Viovy, N. et al. (2005):
Europe-Wide Reduction in Primary Productivity Caused by the Heat and
Drought in 2003. Nature. 437, 529–533.
Huxman,
Travis – Smith, M. – Fay, P. et al. (2004): Convergence
across Biomes to a Common Rain-Use Efficiency. Nature. 429, 651–654.
IUCN
Report (1999): IUCN Report on Biological Diversity on Dryland,
Mediterranean, Arid, Semi-Arid, Savanna and Grassland Ecosystems.
In: Fourth Meeting of the Subsidiary Body on Scientific, Technical
and Technological Advice /Montreal, Canada/.
Reeder,
Jean D. – Schuman, Gerald E. (2002): Influence of Livestock
Grazing on C Sequestration in Semi-Arid Mixed-Grass and Short-Grass
Rangelands. Environmental Pollution. 116, 457–463.
Szente
Kálmán – Nagy Z. – Tuba Z. – Fekete
G. (1996): Photosynthesis of Festuca rupicola and Bothriochloa
ischaemum under Degradation and Cutting Pressure in a Semiarid Loess
Grassland. Photosynthetica. 32, 399–407.
Tuba
Zoltán – Nagy Z. – Czóbel Sz. et al.
(2004a): Hazai gyeptársulások funkcionális
ökológiai válaszai, C-körforgalma és
üvegházhatású gázainak mérlege
jelenlegi és jövőbeni várható éghajlati
viszonyok, illetve eltérő használati módok
mellett. AGRO 21 Füzetek. 37, 123–138.
Tuba Zoltán –
Bakonyi G. – Singh, M. K. (2004b): Impacts on Biodiversity. In:
Láng István – Kőmíves
T. – Jolánkai M. (eds.): Pollution Processes In
Agri-Environment. A New Approach. Akaprint Publishers, Budapest,
235–254.
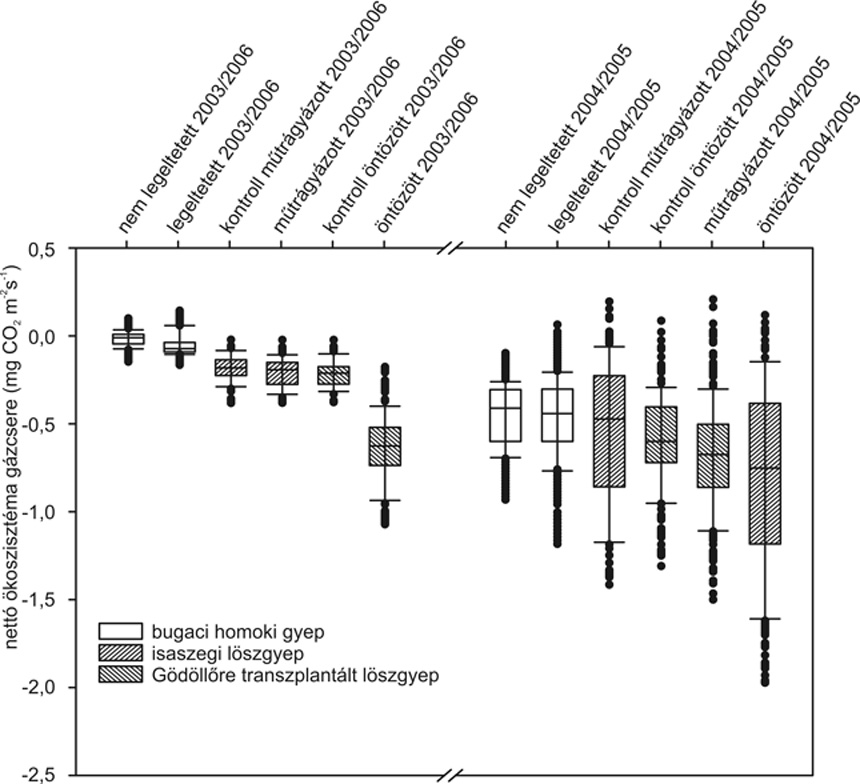
1.
ábra • Nyílt kamrás technikával
mért maximális fotoszintetikus ráta évek
közötti variabilitása megváltoztatott
kezelésű
homoki száraz legelőn
(Cynodonti-Festucetum pseudovinae) és löszpusztaréten
(Salvio nemorosae – Festucetum rupicolae), eltérő
klimatikus viszonyok között. A homoki gyep CO2-megkötése
az optimális vízellátottságú és
fenológiai fázisú évben a vizsgált
kontroll löszgyepekhez hasonló volt. Az extrém
száraz 2003-as év a szünbotanikai viszonyokat és
fiziológiai aktivitást tekintve egyaránt nagyobb
mértékben stresszelte a kevésbé
strukturált, de humuszban gazdag homoki gyepet, mint a
löszgyepet
<--
Vissza a 2007/10 szám tartalomjegyzékére
<--
Vissza a Magyar Tudomány honlapra
[Információk]
[Tartalom] [Akaprint
Kft.]