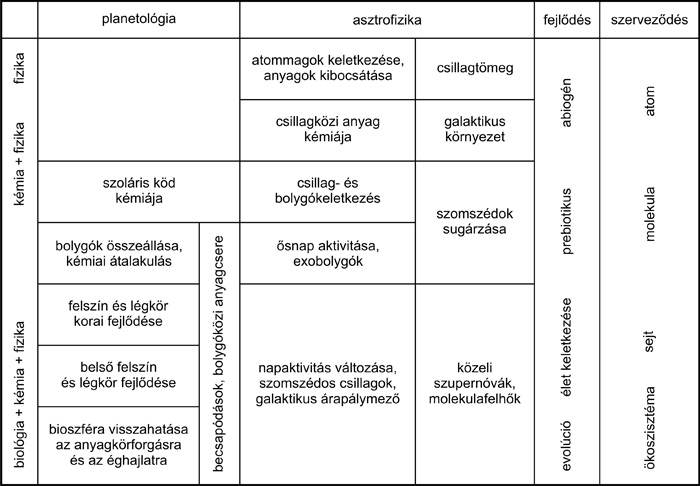
|
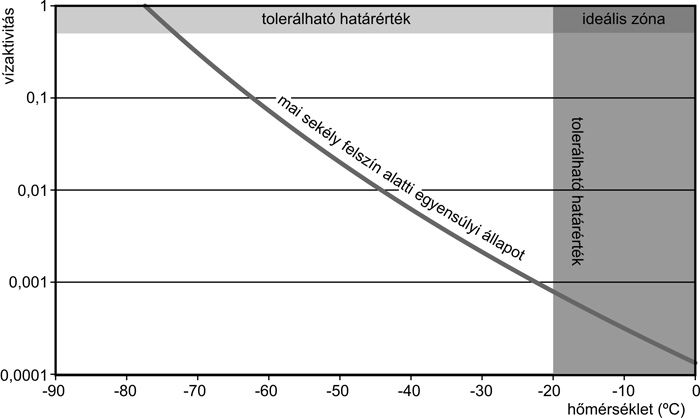
csökken, jégben -40 °C-on
például 0,67. Az ellenálló földi élőlényeknek minimálisan kb. 0,6
körüli vízaktivitás szükséges, ekkor főleg légköri vízpárából fedezik
igényüket. A Marson sajnos (főleg nagy hidegben) elég magas az értéke.
Víz előfordulása: a
felszínen, illetve kis mélységben: vizet a jégsapkákon kívül két
alacsony szélességű foltban, az északi pólussapka körüli
dűnemezőben, a sarkvidéken a jégsapka környezetében
néhány cm-rel a felszín alatt, valamint néhány sziklagleccser
belsejében sikerült eddig kimutatni.
Kedvezőtlen viszonyok: a fentiek
együttesen nem kedveznek a víz megjelenésének. Bár a megolvadáshoz
szükséges hőmérséklet előfordul a Marson, de a szárazság miatt a jég
elszublimál az olvadáspont elérése előtt. Kérdés, hogy vannak-e a
Marson olyan helyszínek/viszonyok, amelyeknél elérhető víz és
megfelelő hőmérséklet együttesen van jelen
(4. ábra).
Alacsony hőmérséklet (Kuti, 2009):
a földi élőlényeknek legalább -20 °C kell a metabolizmushoz, amely
alkalmanként előáll a Mars felszínén, de ennél alacsonyabb
hőmérsékleten is már elszublimálhat a jég. Ezért a vízmolekulákat
erősen kötő anyagok szükségesek, amely akár egy képzeletbeli élőlény
belseje is lehet. Ilyenek különböző erősen higroszkópos tulajdonságú
anyagok, de a diffúziót lassító felszíni portakaró is korlátozhatja a
vízmolekulák elszökését.
Energiaforrások: a felszínen a
napsugárzás, a légköri fotokémiai termékek, a felszín alatt a
kőzet−víz reakciók, főleg a felszabaduló hidrogén és a geotermikus
hővel kapcsolatos reakciók jöhetnek szóba energiaforrásként.
Elképzelhető, hogy az éghajlati
változások (Kereszturi, 2007) nyomán átmenetileg a közelmúltban is
megjelent a víz a felszínen. A tengelyferdeség változásával a sarki
jégsapkák anyaga időnként alacsonyabb szélességre vándorol, majd
visszatér a pólusokra. Eközben nemegyensúlyi állapotok lépnek fel, és
a jég megolvadhat. Talán ezzel kapcsolatos, hogy a Phoenix szonda az
északi sarkvidéken néhány centiméter mélyen karbonátokat,
agyagásványokat és olyan sókat talált, amelyek folyékony vízből
válhattak ki. Az elmúlt millió években több olyan időszak is lehetett,
amikor itt vékony vízfilm borít-hatta a regolit szemcséit, és kémiai
változásokat generált. Egyes megfigyelések és elméleti modellek
alapján az sem zárható ki, hogy mikroszkopikus méretskálán napjainkban
is megjelenhet a víz (Möhlmann, 2004). A Mars tehát érdekes és a mai
technológiával vizsgálható asztrobiológiai célpont.
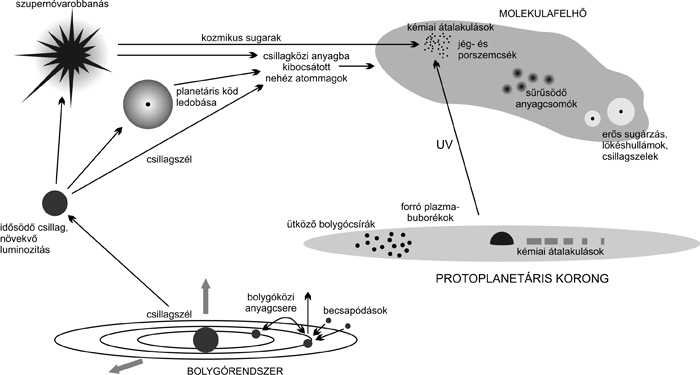
Földi ősleves, kozmikus fűszerek
A földi élet keletkezését megelőző prebiotikus folyamatokat is
befolyásolták kozmikus tényezők. Stanley Miller kísérlete óta tudjuk,
hogy egyszerű szerves anyagok abiogén úton is keletkezhetnek
villámoktól, ultraibolya sugárzástól. A fontos összetevők légköri
keletkezésére a Titan mutat érdekes példát. A korai Földön is
elképzelhető hasonló, az ultraibolya napsugárzástól képződött
szmogréteg, ahol hosszú molekulaláncú, szén alapú anyagok
keletkeztek. De szerves összetevők a kőzetek repedéseit átjáró forró
vizes oldatoktól és a becsapódó testek keltette légköri lökéshullámok
hatására is létrejönnek − mindezek felett pedig magukban a
meteoritokban is érkeztek.
A kondrit meteoritok
szülőégitestjeinek repedéseiben víz cirkulálhatott, kémiai reakciókat
kiváltva. Ezek a meteoritok laza szerkezetük miatt a légkörben néha
szétporladtak, és hamar lelassultak, aminosavakkal és építőköveikkel
bombázva az ősi Földet. Az EET 92042 és GRA 095577 jelű meteoritban
az aminosavak és a fehérjék koncentrációja a más meteoritoknál
jellemző max. 15 ppm helyett 180, ill. 249 ppm volt. A szenes
kondritok anyagának 1–3%-a is lehet szén alapú összetevő, amelynek
harmadát sokgyűrűs aromás szénhidrogének (PAH-ok) tehetik ki.
A Murchison-meteorit például közel
hetvenféle aminosavat tartalmaz, közülük csak hatot használnak a
földi élőlények − emellett az RNS egyik építőköve, egy nukleotidbázis
is megtalálható az anyagában. A Tanzániában 1938-ban hullott Ivuna
nevű, 705 grammos meteoritban pedig az aminosavak közül a prebiotikus
fejlődésben fontos b-alanint és a glicint azonosították. Mindent
összevetve, a becsapódások és a fent említett más folyamatok
együttesen évente közel 108−1010 tonna szerves anyagot termeltek
bolygónk első százmillió éve során. A becsapódó üstökösmagokban is
jöhetett vízjég és szerves anyag, utóbbiak közül a glicint
azonosították a Wild-2 üstökös magjában.
Ugyanakkor az ősi becsapódások
kellemetlenek is lehettek. A 300−400 km-es objektumok ütközései akár a
teljes világtengert elpárologtatták. Ettől forró vízpáralégkör és erős
üvegházhatás alakult ki, és maradt fent ezer évekig. Ilyen
becsapódásokra legutóbb négymilliárd évvel ezelőtt kerülhetett sor.
Extrém élőlények és földi analógiák
Az extrém hidegben vagy melegben, sós, illetve
savas, avagy lúgos környezetben, akár több km mélységig előforduló
extremofilok mint földi analógiák vizsgálata sok érdekes eredményt
hozott. Tűréshatáraikat többféle állapotban is vizsgálhatjuk: 1.
amikor az élőlények szaporodni is képesek; 2. amikor csak anyagcserére
képesek; 3. amikor tetszhalott állapotban inaktívak, de megfelelő
viszonyok esetén még életképesek; és elkülöníthető, amikor már sosem
lesznek életképesek (Gánti, 1971). A jelenleg ismert határokról és
néhány példa taxonról az
1. táblázat
nyújt áttekintést.
Fentiek alapján bolygónkon a forró
vulkáni központokat és a leghidegebb területeket kivéve sok helyen
megélnek egyes extremofilek. A felszín alatt több km mélységig
előfordulnak, amelyek kemoszintézissel, napfény nélkül élnek, és a
kőzet−víz kölcsönhatásokkal keletkező hidrogént, hidrogénszulfidot,
metánt és szénhidrogént használnak fel.
A tetszhalott állapot időtartamát
az összetevők lebomlásának sebessége erősen befolyásolja. Az alacsony
hőmérséklet a lassú kémiai reakciók miatt segít ebben, a felszín
alatti helyzet pedig a kozmikus sugárzástól véd. Kiszáradt állapotban
a sejt citoplazmájában lévő kevés vízmolekulából az ultraibolya
sugárzástól kevesebb reakcióképes és roncsoló gyök keletkezik. A
túléléshez ideális a sporuláció, amikor egy endospórát alkot a sejt,
és ez extrém viszonyokat is túlél, majd megfelelő körülmények közé
kerülve éled újra. A tetszhalott állapotban kibírt időszakra már sok
éve elfogadják a két–négymillió évet, megbízhatónak tekintik a
huszonöt–harmincmillió éves Bacillus sphaericus példányainak
újraéledését is (Cano − Borucki, 1995). Vita tárgyát képezi az eddigi
rekorder, a 250 millió éves 2-9-3 jelű baktériumfaj, amely perm korú
sókristályokból került elő, és egyes szakemberek szerint életképesnek
bizonyult.
Az extremofilek és élőhelyeik
(sivatag, permafroszt, jégtakaró, magashegy, vulkáni környezet)
tanulmányozása támpontot adhat a Földön kívüli élet kutatásához.
Népszerűek például a vastartalomtól vöröses felszínű antarktiszi
szárazvölgyek, ahol a talaj felső rétegében alkalmanként sós oldatok
mozognak, illetve válnak ki. Ugyanitt a Marson megfigyelhető
sárfolyásokhoz hasonló alakzatok is vizsgálhatók. Az antarktiszi
völgyekben ritka a vízfolyás, de a kierodált mélyedés alatt, a
szemcsék között tartósan áramlik az olvadékvíz. A kis völgyben mint
domborzati csapdában dér és hó rakódhat le, amely alkalmanként
megolvadhat.
Sivatagokban nedvességcsapdaként
szolgál a kőzetek málladéktakarójának szemcséi közötti tér. Sivatagi
máznak általában a legkülső, vékonyabb bevonatot nevezik, míg ez alatt
több mm mélységig szintén lehetnek élőlények a mállási kéregben,
amelyet kriptobiotikus kéregnek neveznek (Pócs, 2009). Az
Atacama-sivatagból származó mázakban ATP-molekulák alapján
baktériumokat mutattak ki a legszárazabb vidékeken is. A máz alatt
található 2–4 mm vastag mállási kéreg a hőingástól, a kiszáradástól,
az erős sugárzásoktól védi az élőlényeket − de a fotoszintézishez elég
fényt enged be. A fénymennyiség 2–4 mm mélységig is elegendő az
egyszerű élőlényeknek. Bolygónkon mesterségesen is teremthetünk olyan
viszonyokat, amelyek a marsbéliekhez hasonlítanak. Az ún.
Mars-szimulációs kamrákban alkalmanként meglepően ellenállóknak
mutatkoztak egyes élőlények (Horváth et al., 2006).
A pánspóra elmélet
A pánspóra és pánspermia elméletek szerint élőlények a világűrbe is
kijuthatnak, és ha tetszhalott állapotban túlélik az ott uralkodó
körülményeket, megfelelő viszonyok közé kerülve ismét életre
kelhetnek, élettel „fertőzve meg” egy másik égitestet. Egy nagy
becsapódás a felszínközeli kőzeteket úgy lövi ki, hogy bennük az
ellenálló mikrobák kevéssé roncsolódnak. Svante Arrhenius 1908-ban
vetette fel, hogy a földi globális mágneses tér segítségével is
kerülhetnek baktériumok az űrbe. Az egysejtűeket a felszínről
zivatarok elektromos jelenségei és szelek juttathatják fel 10−50 km
magasba. Itt szaporodnak, és alkalmazkodhatnak az erősebb sugárzáshoz,
kisebb légnyomáshoz és hideghez.
Az egysejtűek felületén megtapadó
töltések a mágneses térrel kölcsönhatnak, és ha a felfelé mutató erő
meghaladja a gravitációs erőt, akár több 100 kilométeres magasságba is
emelkedhetnek a baktériumok. Innen aztán a magnetoszféra
segítségével, például az ún. magnetoszferikus buborékok erővonalaihoz
tapadva távolodhatnak el bolygónktól.
Az egyszerű élőlények tetszhalott
állapotban, rövid idő alatt nem feltétlenül szenvednek el akkora
sugárterhelést az űrben, hogy többé már ne legyenek életképesek.
Hosszabb időt pedig megfelelő sugárvédő réteg segítségével, pl. egy
kőzet belsejében vészelhetnek át. Ekkor viszont már a kődarab saját
radioaktivitása a korlátozó tényező. Végül a légköri belépést és
felizzást kell „túlélnie” az inaktív élőlénynek, amelyre szerencsés
esetben van esély: a meteorikus testnek csak a külső rétege melegszik
át, belül hűvös marad. A nagy sebességű becsapódás pedig a légkörben
bekövetkező szétdarabolódás esetén kerülhető el.
Egy adott bolygórendszeren belül
sokkal nagyobb az esély az élet ilyen vándorlására, mint hogy a
kirepült test egy másik csillag körüli planétán landoljon. A Chicxulub
becsapódás alkalmával például kb. 109−1010 t
anyag repült ki a Földről, amelyből a hozzánk száz fényévnél közelebbi
csillagok környezetébe csak gramm nagyságrendű mennyiség juthatott
el. Jelenleg évente tonnányi nagyságrendű anyag hagyhatja el a
Naprendszert.
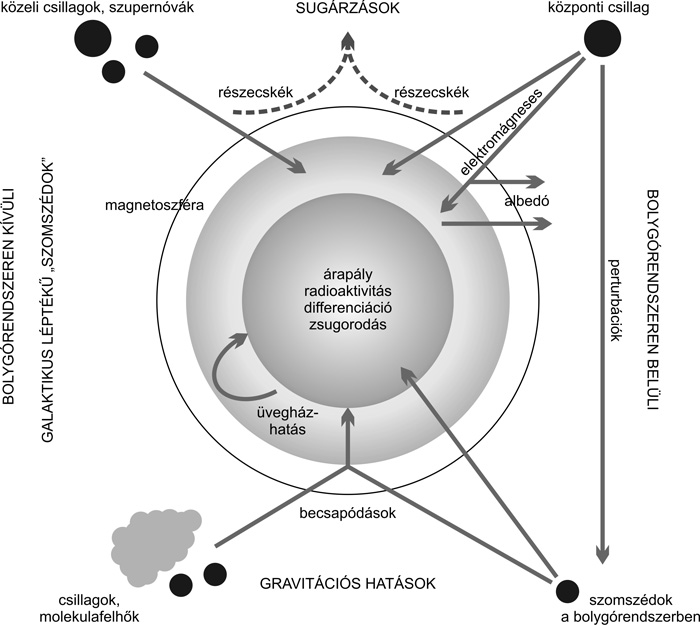
Kozmikus hatások bolygónkra
A bolygórendszeren belüli hatások közül legfonto-sabbak a
becsapódások. Ezek a krátert kialakító és ásványokat átalakító hatásuk
(Mihályi et al., 2008); elfújhatják a légkört, megváltoztathatják az
éghajlatot (például egyes földi kihalások esetében), és átmenetileg
lokálisan megemelhetik a hőmérsékletet. Szerves anyagokat
juttathatnak a felszínre, és akár élőlények égitestek közötti
vándorlásában is közreműködhetnek. A Naprendszer korai időszakában a
Jupiter és a Szaturnusz sok apró égitestet szórt ki, enélkül sokkal
több becsapódás történt volna a Földön is. Az árapályhatástól pedig
változhat a belsőben felszabaduló energiamennyiség,
ami jeget olvaszthat (például Europa) vagy vulkáni aktivitást okozhat
(például Io).
A légkört érő kozmikus sugárzás
(nagyenergiájú atommagok) elősegítik az aeroszolok és a felhők
képződését. Erős napaktivitáskor pedig az intenzívebb napszél
kevesebb töltött részecskét enged a Föld légkörébe, amitől gyengülnie
kellene a felhőképződésnek. Az elméletileg várható kapcsolatot
azonban mérésekkel egyelőre nem sikerült kimutatni. A bolygók
fejlődésére ható külső tényezőket az
5. ábra
foglalja össze.
A lakhatóság fogalma
Az asztrobiológiában régóta használt, gyengén definiált fogalom a
lakhatóság. Szűkebb értelemben azt jelenti, hogy egy adott környezet
biztosítja-e a földihez hasonló élet kialakulásának és
fennmara-dásának lehetőségét. Utóbbit numerikusan jellemezni ma még
nem lehet, ezért egyszerűbb közelítésére a lakhatósági zónákat
használják. Ezek azon térségek egy csillag körül, ahol egy Földhöz
hasonló bolygó felszínén stabilan létezhet folyékony víz. Belső
határán a víz a tropopauzán keresztül a felsőlégkörbe, onnan pedig az
űrbe szökhet. Külső határát nagyjából a légköri szén-dioxid kifagyása
jelenti. A lakhatóság szempontjából fontos még, hogy a csillag
sugárzása stabil legyen. A sok flert produkáló égitestek időnként
lenyomják a bolygólégkörökben az ionopauzát, amitől gyorsul a
légkörvesztés.
Dinamikai szempontból
elkülöníthetők a Nap-rendszerünkben megfigyelt, óriásbolygókon belüli
lakhatósági zóna, a forró Jupiter típusú exobolygók-nál az
óriásbolygóknál távolabb lévő zóna, és amikor maga az óriásbolygó van
a zónában − itt az égitest holdjai lehetnek érdekesek. A lakhatósági
zónákkal kapcsolatban probléma, hogy modelljeik kevés tényezőt
vesznek figyelembe. Emellett nem ismert eléggé a szén-dioxid-felhők
üvegházhatása, és a vulkáni aktivitás fűtőhatásával sem számolnak. Az
élet kialakulásához nem biztos, hogy felszíni víz kell – a felszín
alatt pedig a belső hő vagy árapály-hatás sok helyen olvaszthatja meg
a jeget a csillagtól távol is.
Elméletileg egy csillag galaktikus
környezete is hat bolygóján az élet fejlődésére, de itt még
bizonytalanabb a helyzet. A porban szegény csillagközi zónákban
kevesebb szemcse juthat egy bolygórendszer belsejébe és a bolygók
légkörébe, ami elméletileg csökkentheti a kondenzációs magvak számát
és a felhőborítottságot.
A galaxisok központi fekete
lyukainak környéke vagy a csillagkeletkezési régiókban bekövetkező
gyakori szupernóva-robbanások sugárzásai is kellemetlenek. A
feltételezések alapján extrém nagy tömegű csillagok életének végén
bekövetkező hipernóva-robbanások vagy összeolvadó neutroncsillagok
kiváltotta gammavillanások hatása is jelentős, amelyek fotokémiai
szmogréteget hozhatnak létre. Megnövelik a légköri nitrogén-oxidok és
csökkentik az ózon arányát. Az ezzel kapcsolatban fellépő savas esők a
talajban növelik a nitrátok mennyiségét, aminek kedvező hatása is
lehet. Mivel a csillagkeletkezés a spirálkarokban jellemző, ezért
merült fel a korotációs zóna mint ideális térség lehetősége, ahol a
Nap is megtalálható. Itt az égitestek keringési sebessége a
spirálkarok körbefordulási sebességéhez közeli, azaz ritkán
keresztezik azokat és az ott található csillagkeletkezési
tartományokat.
Hazai kutatások
Az exobolygókkal kapcsolatos hazai kutatások közül kiemelkedik a
Hungarian Automated Telescope (HATNet) amerikai−magyar távcsőhálózat
(Bakos et al., 2004), amely 2009 februárjáig tizenegy fedési
exobolygót talált. A magyar ötlet alapján készült távirányítású
robotteleszkóp-rendszert itthon tervezte és építette Sári Pál, Papp
István és Lázár József (Magyar Csillagászati Egyesület) Bakos Gáspár
(korábban MTA KTM CSKI, ma Harvard-Smithsonian Asztrofizikai Központ)
vezetésével. A bolygókeresés szoftveres oldalára Kovács Géza (MTA KTM
CSKI) dolgozott ki hatékony módszereket. Exobolygókkal is kapcsolatos
a Magyar Asztroszeizmológiai Csoport munkája, amelynek eredményeit a
V391 Pegasi b jelű exobolygó felfedezéséhez használták fel. A Szegedi
Tudományegyetem és a Szegedi Csillagvizsgáló munkatársai (Simon et al.
2007) az exobolygókkal és holdjaikkal kapcsolatos modellezésen
dolgoznak. Az ELTE Csillagászati Tanszékén pedig a távoli planéták
pályaelemeinek stabilitásával kapcsolatos számításokat végeznek Érdi
Bálint vezetésével (Érdi − Sándor, 2005).
A Collegium Budapest Institute for
Advanced Study intézetben az Európai Űrügynökség és a Magyar
Űrkutatási Iroda támogatásával működő Mars Asztrobiológia
Kutatócsoport a Mars felszíni viszonyait, a víz időszakos megjelenési
lehetőségét és potenciális földi analógiákat, extremofil
szervezeteket vizsgál (Szathmáry et al., 2007). A szerves anyagok
ultraibolya sugárzásra adott reakcióját tanulmányozzák az MTA–SE
Biofizikai Kutatólaboratóriumában. Rontó Györgyi és Bérces Attila
vezetésével hazánk is részt vesz a világűri kitettségi vizsgálatokban,
ahol a kozmikus sugárzás hatását tanulmányozzák, részben a
pánspóra-elmélet szempontjából.
Az asztrobiológiai ismeretek és
módszerek az egyetemi oktatásban is hasznosak a téma érdekessége és
az eltérő tudományterületek közötti kapcsolódási pontok miatt.
Asztrobiológiai kurzusok hangzottak el az ELTE Csillagászati Tanszéke
szervezésében Almár Iván és Illés Erzsébet vezetésével 1997-ben,
később a Szegedi Tudományegyetemen, és az ELTE Természetföldrajzi
Tanszékén. A nagyközönséget célozza meg az Origo online Origo
asztrobiológia kurzus című cikksorozata, amelynek részeit alkalmanként
10–15 ezer olvasó böngészi (Simon − Kereszturi 2009).
Az eltérő tudományterületek és a
műszaki szakemberek kooperációjára ideális keretet nyújt az
asztrobiológa. A fejlesztés alatt álló űrteleszkópok tervezésénél
geológusok, biológusok, meteorológusok és vegyészek mérnökökkel
együtt dolgoznak, hogy a berendezések képesek legyenek a légköri
összetétel meghatározásán túl felhőket, víztükröt, éghajlati
jellemzőket kimutatni az exobolygókon. A színképekben talán a
szárazföldeket sokáig borító, baktériumokból álló bevonathoz hasonlót,
esetleg klorofilt is azonosíthatnak. Eközben a Mars-szimulációs
kamrából nyert eredményeket az antarktiszi McMurdo-szárazvölgyek
megfigyeléseivel szintetizálva tervezik a következő Mars-szondákat,
amelyek életnyomokat kereső műszereit az Atacama-siva-tagban is
tesztelik, szintén asztrobiológiai projektek keretében.
Kulcsszavak: asztrobiológia, Földön kívüli élet, Mars,
Europa, Titan
IRODALOM
Almár Iván (1999): A SETI
szépsége. Vince, Budapest
Bakos Gáspár Á. − Noyes,
R. W. − Kovács G. − Stanek, K. Z. − Sasselov, D. D. − Domsa I. (2004):
Wide-field Millimagnitude Photometry with HAT: A Tool for Extra-Solar
Planet Detection. Publications of the Astronomical Society of the
Pacific –PASP. 116, 266–277.
http://www.journals.uchicago.edu/doi/pdf/ 10.1086/382735
Cano, Raul J. − Borucki,
Monica K. (1995): Revival and Identification of Bacterial Spores in
25- to 40-million-year-old Dominican Amber. Science. 268, 5213,
1060–1064.
Érdi Bálint − Sándor Zsolt
(2005): Stability of Co-Orbital Motion in Exoplanetary Systems.
Celestial Mechanics and Dynamical Astronomy. 92, 1–3, 113–121.
Gánti Tibor (1971): Az
élet princípiuma, Gondolat, Budapest
Mihályi Krisztián – Gucsik
A. – Szabó J. (2008): Drainage Patterns of Terrestrial Complex
Meteorite Craters: A Hydrogeological Overview. 39th Lunar and
Planetary Science Conference #1200.
Gyenizse Péter (2008):
Planetomorfológia. In: Lóczy Dénes (szerk.): Geomorfológia II. Dialóg
Campus, Budapest–Pécs, 305–362.
Horváth András − Gánti T.
− Bérczi Sz. − Pócs T. − Kereszturi Á. − Sik A. (2006): Marsi
dűnefoltok: az élet lehetősége a Marson? Magyar Tudomány. 11,
1357–1375.
Kereszturi Ákos (2007): Éghajlatváltozás a Marson I–II. Légkör. 52, 2,
12–17., 52, 3, 6–9.
Kereszturi Ákos − Simon
Tamás (2005): Asztrobiológia. In: Mizser Attila – Taracsák G. –
Szabados L. (szerk.): Meteor csillagászati évkönyv. Magyar
Csillagászati Egyesület, Budapest, 190–218.
Kuti Adrienn (2009):
Thermal Behavior of Dokka Crater and Its Surroundings in the North
Polar Region of Mars. 40th Lunar and Planetary Science Conference
#1006.
Mizser Attila − Kereszturi
Ákos (2003): The Astrobiology Matrix and the “Drake Matrix” in
Education. 34th Lunar and Planetary Science Conference Abstract 1114.
Möhlmann, Dietrich T. F.
(2004): Water in the Upper Martian Surface at Mid- and Low-latitudes:
Presence, State, and Consequences. Icarus. 168, 318–323.
Pócs Tamás (2009):
Cyanobacterial crust types, as strategies for survival in extreme
habitats. Acta Botanica Hungarica. 51, 1–2, 147–178.
http://www.colbud.hu/esa/publications/
Cyanobacterial_crust_types.pdf
Simon Tamás − Kereszturi
Ákos. (2009): Online Astrobiology Course in Hungary. 40th Lunar and
Planetary Science Conference abstract 1048.
Simon Attila − Szatmáry K.
− Szabó Gy. M. (2007): Determination of the Size, Mass, and Density of
”Exomoons” from Photometric Transit Timing Variations. Astronomy &
Astrophysics. 470, 727–731.
Szathmáry Eörs − Gánti T.
− Pócs T. − Horváth A. − Kereszturi A. − Berczi Sz. − Sik A. (2007):
Life in the Dark Dune Spots of Mars: A Testable Hypothesis. In:
Pudritz, Ralph – Higgs, P. – Stone, J. (eds.): Planetary Systems and
the Origin of Life. Cambridge Astrobiology Series III., Cambridge
University Press
|