|
Genetika
Az immunglobulin-szerű KIR-gének
Az NK-gének emberben részben a 19. kromoszómán, az úgynevezett
leukocita receptor területen helyezkednek el. Fontos jeltovábbító
molekulák génjei is találhatók itt, melyeket az NK-receptorok is
használnak működésük során. A gének öröklődéséből következik, hogy
mindenki számos olyan MHC ellen irányuló KIR-receptorral bír, amelynek
liganduma nem is fordul elő a saját szervezetében, ám minden
NK-sejtnek van a saját HLA-ra specifikus receptora. Ez azt jelenti,
hogy őssejtátültetés esetén a KIR-receptorokkal szembeni (NK)
összeférhetetlenség valószínűsége jelentősen emelkedik.
Egy emberben legalább tizenöt különböző
KIR-receptorforma ismert, amelynek több száz örökölhető variációja
fordul elő (Barten et al., 2001). Ezt a magasfokú eltérést a genetikai
okok magyarázzák.
Lektinkötő receptorgének
A receptorok a 12. kromoszóma régióban kódoltak. Kálciumion-függő, a
sejtfalon áthatoló fehérjék, amelyeknek a sejten kívüli része köti meg
a lektint, amely valamennyi receptor esetében azonos, és
glikoproteinhez kötődik. Gátló vagy aktiváló tulajdonságot hordoznak,
és ősi molekulákat mint ligandumokat ismernek fel (HLA-E-F).
A KIR-család a 19-es, a lektin receptorcsalád pedig
a 12-es kromoszómán helyezkedik el. A receptorok és a ligandumok is
egymástól függetlenül öröklődnek. Az MHC- (HLA) génektől eltérő
öröklődés azt jelenti, hogy HLA-ban egyező őssejtdonorok az NK-sejtek
KIR-receptoraiban még a nem egypetéjű ikrek esetében is eltérőek
lehetnek (Wilson et al., 1997).
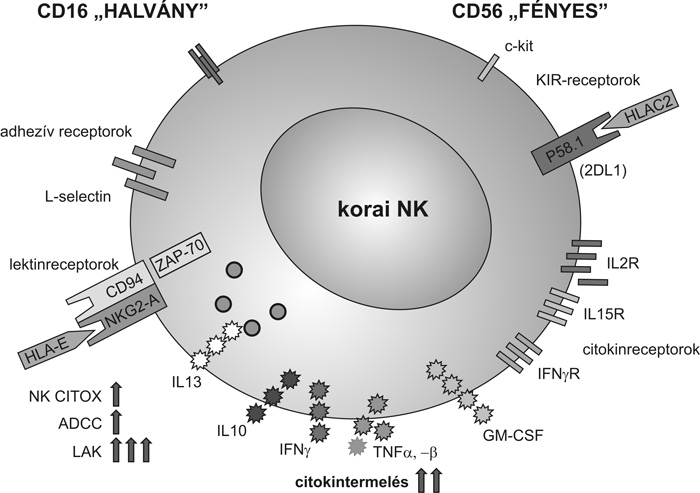
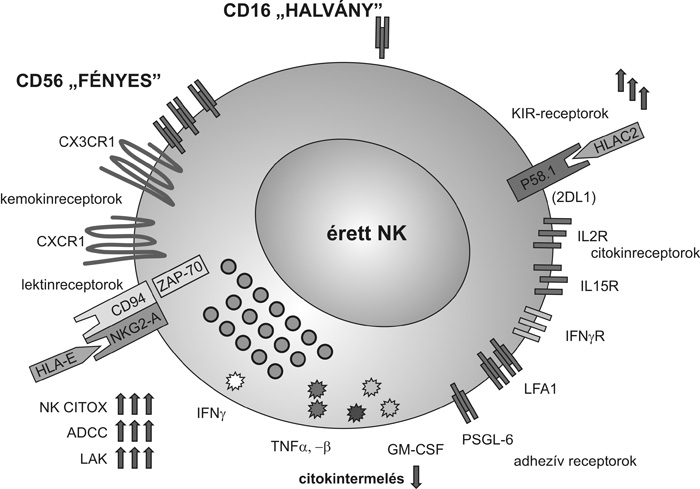
NK-sejttípusok
Az utóbbi években lehetőség nyílott az NK-sejtek KIR receptorainak
standard genetikai módszerekkel (például génamplifikáció-PCR) történő
meghatározására (Steffens et al., 1998). Ennek fontosságát az újabb
klinikai adatok tükrében ítélhetjük meg.
Érdekesek azok a microarray (chip) vizsgálatok,
amelyek arról számolnak be, hogy bizonyos NK-gének csak a korai, mások
a késői NK-sejtekben fejeződnek ki, és a gének egy része mindkét
sejtcsaládban aktív (Wilk et al., 2008).
Újabban a gének olyan kombinációjáról számoltak be,
amelyek együttes mintázata csak az NK-sejtekre jellemző, és
megkülönbözteti az NK-sejteket az immunrendszer további sejtjeitől
(Walzer et al., 2007).
Klinikai megfigyelések
Mind jobban elterjedt, hogy rosszindulatú folyamatok (leukémiák,
limfómák) és egyes immunhiányos állapotok gyógyítására emberi
őssejtátültetést végeznek. Az ennek előkészítésére alkalmazott
eljárások (gyógyszeres kezelés vagy egész test besugárzás) szinte
teljesen elpusztítják a befogadó szervezet saját immun- és vérképző
sejtjeit. Az őssejtátültetéseket (csontvelő-transzplantáció) követő
regenerációban a periférián megjelenő első sejtcsalád az NK-sejtek.
Az eddigi klinikai megfigyelések azt bizonyítják,
hogy az őssejtdonor és a befogadó (recipiens) szervezet közötti
NK-egyezés egyértelműen befolyásolhatja a beültetett sejteknek a
gazdaszervezet ellen meginduló immunreakcióját, amely az életet
veszélyeztető (ún. graft versus host – GVH) betegség formájában
nyilvánul meg. További megfigyelések kellenek azonban ahhoz, hogy
ennek a viszonynak a részleteit is megismerhessük. Ésszerű tehát, hogy
az NK-sejtek receptorainak genetikai módszerekkel történő
meghatározása teret nyerjen; és az eddigi eredményekből bevezetésre
kerüljenek azok az eljárások, amelyek a betegek javát szolgálhatják.
Ismeretes, hogy a fehérvérűségek gyógyításának előkészítésénél a
lehetséges maximális immunoszuppressziós (besugárzásos) kezelés sem
pusztítja el az összes daganatos sejtet. A megbúvó maradék
rosszindulatú sejtből a betegség kiújulhat. Szükséges ezért, hogy az
átültetett őssejtekből kinövő új T-limfocita sejtcsaládok felismerjék
a megbúvó sejteket, és ellenük immunreakciót (GVH) indítsanak meg. A
reakció erős formája a GVH-betegség (GVHD), amely rossz esetben a
betegek halálát okozhatja. A befogadó szervezettel reagáló
NK-sejtekről a jelenlegi klinikai megfigyelések alapján
valószínűsíthető, hogy a megbúvó daganatsejteket elpusztítják.
A KIR-receptorok jelentőségét a legújabb fontos
eredmények húzzák alá; 202 HLA-azonos testvér közötti
őssejttranszplantált beteg klinikai adatainak elemzésével
bizonyították, hogy a transzplantáció eredményessége függ a donor és
recipiens KIR genotípusától. Aktiváló típusú KIR-receptor jelenléte a
donorsejteken emelte az immunreaktivitást, ugyanakkor a befogadó beteg
gátló KIR-receptorai az elfogadást segítették elő. További kiterjedt
vizsgálatok és klinikai megfigyelések szükségesek a fenti ígéretes
eredmények megerősítésére és az aktiváló/gátló KIR receptorok
immungenetikai tesztelésére, amelyek jelentősen megváltoztathatják az
őssejt-transzplantációs terápiás eljárásban eddig követett
kiválasztási stratégiánkat. Joel Y. Sun és munkatársai (2005) azt
észlelték, hogy a nemrokon őssejtátültetést követő GVHD erősségét
növelte, ha a leukémiás betegek több gátló KIR-receptorral bírtak,
mint a donorok, és/vagy a donoroknak több aktiváló receptoruk volt,
mint a betegeknek. Japán szerzők 379 vastagbéldaganatos beteg
vizsgálatával megállapították, hogy a magas NK-aktivitás védő hatású e
daganattípusban (Furue et al., 2008).
Terhesség és NK-sejtek
A méhbe beágyazódó petesejt, illetve embrió genetikai okok miatt
immunológiailag „félig idegen”, hiszen az apai géneket is kifejezi. A
szervezet azonban különböző utakon és változatos mechanizmusokkal
védi az embrió zavartalan fejlődését. A korai szakaszban egy speciális
NK-sejtcsalád működik ennek érdekében. Korábbi vizsgálatok kimutatták,
hogy a terhesség első három hónapjában a méhben speciális
NK-sejtcsalád alkotja a sejtek mintegy 70–90%-át. Ezek az ún.
uterus natural killer sejtek (uNK) a méh átalakult
nyálkahártyájában találhatók, és hormonális szabályozás alatt állnak.
Aktivált NK-sejteknek tekinthetők, amelyeknek sejtfelszíni jellemzői
is eltérőek, és a korai (embrionális) NK-sejtekre emlékeztetnek.
Az embrióburkokon az ősi HLA-molekula (HLA-G és
-E) fejeződik ki. A G-molekula gátolja, hogy az uNK-sejtek felismerjék
a szemi-allogén trofoblaszt sejteket, és így elősegíti az embrió
beágyazódását és fejlődését (O’Connor et al, 2006). További és újabb
vizsgálatok szerint előtérben álló gátló mechanizmus a HLA-E-molekula
és a megfelelő lektinreceptor kapcsolat, amely fiziológiás terhesség
esetén gátló hatást fejt ki. Mindez ésszerű; ám kellőképpen nem
bizonyított, hogy az uNK-sejtek a „félig idegen” embriót ismerik fel,
és végső soron az első három hónapban a terhesség fennmaradásában a
legfontosabb immunológiai tényezőt jelentik. (Lanier, 1998).
A legújabb felfogás szerint a korai élettani
terhességet mint az uNK-sejtek–trofoblaszt sejtek interakciója
következményeként kiváltott steril gyulladásos reakciójának tekintik,
aminek hatására az immunrendszert stimuláló üzenetközvetítő
molekulákat (citokineket) termelnek. Az egészséges terhességben azok a
citokinek (Th2) játsszák a főszerepet, amelyeket korai típusú
uNK-sejtek termelnek. Ismétlődő vetélésekben az uNK-sejtek késői
típusúak, és főként más (Th1) citokineket termelnek, amelyek további
immunsejtek aktiválásával a magzat korai felszívódását váltanák ki
(Sargent et al, 2007). A valóságos szabályozó mechanizmus ennél az itt
vázolt, egyszerűsített képnél minden bizonnyal bonyolultabb, amit az
idevágó kiterjedt irodalom is jelez.
Fontos kiemelnünk, hogy az uNK-sejtek hormonális
hatás alatt állnak; noha hormonreceptoruk nincsen, mégis követik a női
ciklus ingadozását. Hazai kutatók kimutatták, hogy sárgatesthormon
hatására limfocitákban termelődő gátló faktor található (PIBF –
progeszteron indukált blokkoló faktor), amely citokinközvetített
mechanizmus révén csökkenti az uNK-sejtek citotoxikus aktivitását,
vetélés ellenes hatást fejtve ki (Szekeres-Barthó et al., 1996;
Anderle et al., 2008)
Mindebből a rövid összefoglalásból is látható, hogy a szabályozás
bonyolult láncolatában az uNK-sejtek fontos szerepet töltenek be,
amelyeknek további szabályozó körei már részben ismertek.
Cikkünkben az emberi természetes ölősejtek legfontosabb jellemzőit,
genetikai-immunbiológiai tulajdonságait foglaltuk össze. Elsősorban
azon területeket tekintettük át, ahol az elmúlt években az új
megismerések klinikai szempontból is számottevőek, illetve azokat,
amelyek felfogásunkat változtatták meg e kevéssé ismert sejtcsaládról.
RÖVID FOGALOMMAGYARÁZAT
receptor • sejtes szerkezeti elem, amely a
neki megfelelő elemmel (ligandummal) kötődve jelet továbbít a sejt
magja felé
KIR • (killer immunglobulinszerű receptor)
az NK-sejtek immunglobulin szerkezethez hasonló felismerő receptorai
ligandum • a receptorhoz specifikusan kötődő
molekula
MHC-I • minden magvas sejten, az egyed
szöveti azonosságát hordozó molekulacsaládok genetikai kódját hordozó
szakasz
(HLA-A-B-C) • az MHC-I által kódolt szöveti
kompatibilitási jegyek (transzplantációs antigének)
T-sejtek • immunológiai válaszképességüket a
magzati életben a csecsemőmirigyben (timusz) elnyerő fehérvérsejtek,
amelyek főként a szerzett immunitást (és az immunológiai emlékezetet)
biztosítják
citokinek • sejtek által termelt
üzenetközvetítő, szabályozó molekulák
GVH-reakció • (graft versus host – GVH) a
beültetett immunkompetens sejteknek a befogadó szervezet elleni
immunreakciója
GVHD • az előző immunreakció következtében
kialakuló súlyos betegségegyüttes
immunszuppresszió • a szervezet
immunválaszának elnyomása (sokszor gyógyító célzattal, gyógyszer,
besugárzás alkalmazásával)
AML • a fehérvérűség (leukémia) egyik
formája, amely a fehérvérsejtek csontvelői előalakjaiból indul ki
uNK • a terhesség első harmadában, a méhben
található különleges NK-sejtek
Kulcsszavak: immunológia, immunválasz, természetes ölősejt,
limfocita, receptor, klinikum, sejtaktiváció
IRODALOM
Anderle, Christine – Hammera, A. – Polgár
B. et al. (2008): Human Trophoblast Cells Express the Immunomodulator
Progesterone-Induced Blocking Factor. Journal of Reproductive
Immunology. 79, 26–36.
Barten, Roland – Torkar, M. – Haude, A. et
al. (2001): Divergent and Convergent Evolution of Nk-Cell Receptors.
Trends in Immunology. 22, 52–57.
Benczúr Miklós – Sármay G. – Laskay T. et
al. (1982): Recognition of Autologus and Allogeneic Lymphocytes and
Tumor Cells by Human Natural Killer Cells. Molecular Immunology. 19,
1331–1340.
Benczúr Miklós (2006): Emberi Nk sejtek
allogén felismerő képessége és funkciója az őssejtek
transzplantációjában. Hematológia Transzfuziológia. 39, 5–9.
Cooper, Megan A. – Fehniger, T. A. –
Caligiuri, M. A. (2001): The Biology of Human Natural Killer-Cell
Subsets. Trends in Immunology. 22, 633–640.
Furue, Hiroki – Matsuo, K. et al. (2008):
Decreased Risk of Colorectal Cancer with the High Natural Killer
Activity NKG2D Genotype in Japanese. Carcinogenesis. 29, 316–320.
WEBCÍM >
Lanier, Lewis L. (1998): Nk Cell
Receptors. Annual Review of Immunology. 16: 359–393.
Ljunggren, Hans-Gustaf – Kärre, Klas
(1990): In Search of the ‘Missing Self’: Mhc Molecules and Nk Cell
Recognition. Immunology Today. 11, 237–244.
Maghazachi, Azzam A. (2005):
Compartmentalization of Human Natural Killer Cells Molecular
Immunology. 42, 523–529.
Moretta, Lorenzo – Bottino, C. – Pende, D.
et al. (2005): Human Natural Killer Cells: Molecular Mechanisms
Controlling Nk Cell Activation and Tumor Cell Lysis. Immunology
Letters. 100, 7–13.
O’Connor, Geraldine M. – Hart, O. M –
Gardiner, C. M. (2006): Putting the Natural Killer Cell in Its Place.
Immunology. 117, 1–10.
Sargent, Ian L. – Borzychowski, A. M. –
Redman, W. G. (2007): Nk Cells and Human Pregnancy – An Inflammatory
View. Trends in Immunology. 27. 399–404.
Steffens, U. – Vyas, Y. – Dupont, B. –
Selvakumar, A. (1998): Nucleotide and Amino Acid Sequence Alignment
for Human Killer Cell Inhibitory Receptors (Kir). Tissue Antigens. 51,
4, 398–413.
Sun, Joel Y. – Gaidulis, L. – Dagis, A et
al. (2005): Killer Ig-like Receptor (Kir) Compatibility Plays a Role
in the Prevalence of Acute GVHD in Unrelated Hematopoietic Cell
Transplants for AML. Bone Marrow Transplant. 36, 525–530.
WEBCÍM >
Szekeres-Barthó Júlia – Faust Zs. – Varga L. et al. (1996): The
Immunological Pregnancy Protective Effect of Progesterone Is
Manifested Via Controlling Cytokine Production. American Journal of
Reproductive Immunology. 35, 348–351.
Trinchieri, Giorgio (1984): Biology of
Disease of Human Nk Cells: Biological and Pathological Aspects.
Laboratory Investigation. 50, 489–513.
Vivier, Eric (2006): What Is Natural in
Natural Killer Cells? Immunology Letters. 107, 1–7.
Walzer, Thierry – Jaeger, S. – Chaix, J.
et al. (2007): Natural Killer Cells: From Cd3 Snkp46+ to Post-genomics
Meta-analyses. Current Opinion in Immunology. 19, 365–372.
Wilk, Esther – Kalippke, K – Buyny, S et al. (2008): New Aspects of Nk
Cell Subset Identification and Inference of Nk Cells’ Regulatory
Capacity by Assessing Functional and Genomic Profiles. Immunobiology.
213, 271–283.
Wilson, Michael J. – Torkar, M. – Haude,
A. et al. (1997): Plasticity in the Organization and Sequences of
Human Kir/Ilt Gene Families. Proceedings of the National Academy of
Sciences of the USA. 97, 4778–4784.
Zhang, Cai – Zhang, J. – Wei H. (2005):
Imbalance of Nkg2d and Its Inhibitory Counterparts: How Does Tumor
Escape from Innate Immunity? International Immunopharmacology. 5,
1099–1111.
|