|
|
A Darwin-emlékév kapcsán nem tűnik
haszontalannak a növényvilág evolúciójáról szerzett ismereteinket a
darwini fejlődéselmélet szemszögéből áttekinteni, és állításait a
növényvilág szempontjából értékelni. Az egyik ok, amelyért ezt meg
kell tennünk az, hogy a darwini elmélet alapvetően az állatvilágban
megfigyelt alkalmazkodási jelenségekre épült, holott a földi élet
története szempontjából a kulcsfontosságú szerep a növényvilágé,
amely a földi élő rendszereket működtető kozmikus energiaforrás
egyetlen akceptoraként egyrészt a földi légkört az élő szervezetek
számára élhető közeggé formálta, másrészt szerves tápanyaggal ellátta.
Másként fogalmazva az állatvilág − az embert is beleértve − a
növényvilág vendégeként éli földi életét.
Tanulmányomban szeretnék rávilágítani néhány momentumra, amelyek a
darwini elméletet nemcsak zseniálisnak, hanem vitathatónak is ítélik.
A darwini elmélettel részben az a gond, hogy belőle már Charles Darwin
életében ideológia lett, ami nem mindig kedvező egy tudós számára, és
nem biztos, hogy ez a tény megegyezik a tudós eredeti szándékával.
Kétségtelen, hogy hírneve már életé-ben hatalmas mértékben megnőtt, és
Darwin elméletéből nemcsak darwinizmus lett, hanem neodarwinizmus
is, ami a 20. század második felének jelentős szellemi áramlatává
vált.
Ez utóbbinak képviseletében Richard Dawkinst idézhetjük, Az önző
gén és A vak órásmester című könyvek szerzőjét, aki többek
között ezt írja: „A természet egyedüli órásmesterei a fizika vak
erői, bár ezek nagyon különleges módon fejtik ki hatásukat. Egy valódi
órásmester előrelátóan cselekszik: megtervezi fogaskerekeit és
rugóit, valamint ezek kapcsolatait, miközben egy jövőbeli cél lebeg a
szeme előtt. A természetes szelekciónak, a vak, tudattalan,
automatikus folyamatnak, amit Darwin fedezett fel, és ami mai tudásunk
szerint megmagyarázza az élet összes formájának létezését és
látszólagos célját, semmilyen cél nem lebeg a szeme előtt. Nincs
elméje és nincsenek lelki szemei, és nem tervez előre semmit. Nincs
látomása, előrelátása, egyáltalán semmilyen látása nincs. Mondhatni ő
játssza az órásmester szerepét a természetben, ráadásul ez az
órásmester teljesen vak.” (Dawkins, 1986, 14.)
Ezzel szemben Darwinnak nem állt szándékában az élet összes formáinak,
és különösen nem az élet keletkezésének megmagyarázása, amint maga
írja: „Előre kell bocsátanom, hogy sem a szellemi képességekkel, sem
az élet eredetével nem kívánok foglalkozni” (Darwin, 1958, 331.). Úgy
tűnik, hogy Darwin elméletét olyan elvárásokkal terhelték meg és
teszik ma is, amelyeket nem biztos, hogy el tud viselni. Az sem
szerencsés, ha egy tudományterület problémáinak megoldását egy másik
tudományra hárítják át, amelyben az illető nem szakember. Egy biológus
ne mondja azt, hogy a biológiai folyamatokat a „fizika vak erői”
irányítják. Az élő szervezeteknek mindig is megvoltak az eszközeik,
hogy a fizika „vak erőit” saját javukra fordítsák.
Egy vizsgálható fejlődésmodell: a szukcesszió
Az evolúció megítélésének fő nehézsége, hogy nem tudjuk lejátszani,
mert ez az eseménysor egyszer zajlott le a Föld története során. De
modellként vizsgálhatunk olyan jelenben lejátszódó folyamatokat,
amelyek egyértelműen fejlődési folyamatok, és a szereplői növények.
Ilyen például egy trópusi esőerdő regenerációja egy hurrikán által
letarolt területen, ahol a talajt elvitte az erózió, és a környezetből
indul el a terület újraerdősülése. A Karib-térségben, ahol ilyen
természeti katasztrófák évente megtörténnek, számos alkalommal
vizsgáltak ilyen folyamatokat, magam is vizsgáltam ilyet. Azt látjuk,
hogy a különböző szaporodási stratégiával rendelkező fajok
meghatározott sorrendben váltják egymást a lepusztult termőhelyen, és
hoznak létre egy sekély termőtalajt, amelyen gyorsan növő puhafák
alakítják ki az első erdei közösséget, amely 15–30 év alatt létrehoz
egy 10–15 méter magas koronaszintű állományt. Ezt nevezzük első
homeosztázisnak, vagyis a fejlődés első szakaszának. Ez a közösség
igen nagy mennyiségű, gyorsan bomló szerves anyagot termel, amely
felgyorsítja a talajképződési folyama-tokat, és ezen a megnőtt
tápanyagbázison újabb huszonöt–ötven év alatt kialakul magas fákból
egy második, immár 25–30 m magas koronaszint. Ez a második
homeosztázis, amelynek élettartama már 50–100 év, és amely alatt
megtaláljuk az alsó koronaszintet is. A trópusi esőerdők dinamikáját
kevésbé ismerő szerzők már ezt az együttest valódi, kész trópusi
erdőknek gondolják, és ezért írják tévesen, hogy a trópusi esőerdők
ötven év alatt regenerálódnak. A fejlődésnek ugyanis még nincs vége.
Ezután jelennek meg és nőnek fel a 40–50, esetleg 70 m magasra megnövő
óriásfák, kialakítva a trópusi esőerdő harmadik, felső koronaszintjét.
Ez a harmadik homeosztázis, amely 150–300 év alatt alakul ki, és az
óriásfák koronaszintjében fejlődik ki a liánok és epifiton
szervezetek óriási gazdagsága a bennük élő rovarok, madarak,
emlősállatok hihetet-len változatosságú élővilágával együtt. Ez az a
közösség, amelynek élete egyensúlyban van az éghajlati
körülményekkel, vagyis annyi szerves anyagot termel, amennyit beépít a
szervezetébe, és eközben fenntartja a talaj táperejét. Ezzel szemben
az első két homeosztázis szereplői sokkal több szerves anyagot
termeltek, mint amennyit testükbe beépítettek, és
többlettermelésükkel a következő homeosztázis feltételeit teremtették
meg. Mindebből megállapítható, hogy a szukcesszió egy szakaszos
fejlődési folyamat, amelyben felépítő és egyensúlyi fázisok váltják
egymást. Ebben a folyamatban sem a fizikai, sem a biológiai erők nem
vakon működnek, a folyamat annyira felismerhető momentumokból áll,
hogy ezek ismeretében az erdővagyon nagysága és évi növekménye
kiszámítható, amelyre gazdálkodási terv építhető, és vele mint
megújuló energiaforrással fenntartható módon gazdálkodhatunk. Vagyis:
a természeti erőforrásokkal való kiszámítható és tervezhető
gazdálkodás ténye ellentmond az erőforrások vak és véletlenszerű
keletkezésének.
Segíthetnek-e a molekulák?
Nagy várakozással tekintettünk az utóbbi két évtized molekuláris
genetikai és taxonómiai kutatásaira, amelyek segítettek abban, hogy az
örökítő anyag szerkezetéből, a DNS-molekula bázissorrendjeinek
összehasonlításából megismerjük az evolúció eddig homályos részleteit,
hogy az élő szervezetek hasonlóságára épített fejlődéselméleteket
konkrét genetikai struktúrákkal igazoljuk. Azt vártuk, hogy a
megismert génszekvenciák a lezajlott fejlődés sorrendjébe állítják a
ma ismert növényvilágot, és végre megszületik az egyetlen hiteles
evolúciós törzsfa, amely felrajzolja a növényvilág fejlődéstörténetét
úgy, hogy közben betömi azokat a hézagokat, amelyeket a darwini
elmélet nyitva hagyott. Munkatársaimmal magam is bekapcsolódtam ilyen
kutatásokba.
Ahogyan ez lenni szokott egy új módszer bevezetésekor, a molekuláris
genetikai-taxonómiai kutatások igen sok nyitott kérdést megoldottak.
Az evolúciós törzsfák számos korábbi származástani feltételezését
megerősítették, sok új meglepő eredményt is hoztak, de miközben sok
korábbi tudáshézagot betömtek, nagy számban vetettek fel új
problémákat is. (Borhidi, 2008)
A kladisztikai törzsfák azonnal két meglepetéssel szolgáltak.
Korábban, a morfológiai hasonlóságokra épülő törzsfákon úgy tüntettük
fel a fejlődést, hogy a fejlettebb csoportokat a földtörténet során
korábban megjelent, ismert fejletlenebb csoportokból származtattuk.
Ezzel szemben a mai molekuláris törzsfák elágazásain mindenütt
ismeretlen, feltételezett csoportok szerepelnek, amelyeket még
megnevezni sem tudunk. Vagyis, eljutottunk oda, hogy van egy olyan
konkrét genetikai információkon felépülő evolúciós törzsfánk, amelynek
valamennyi elágazási pontja hipotetikus. Úgy is fogalmazhatnánk, hogy
amennyivel okosabbak lettünk a részleteket illetően, annyival
bizonytalanabbak az egész megítélésében. Mai ismereteink ugyan
kétségtelenül valósabbak, de az összkép homályosabbnak tűnik, mint
volt korábban. A törzsfák másik problematikus eleme, hogy a fejlődés
folyamatát ábrázoló származási vonalak mentén számok szerepelnek,
azon tulajdonságok száma, amelyek a vonal végén szereplő
növénycsoport kizárólagos sajátjai, amelyek a szomszédos, ún.
„testvércsoporttól” megkülönböztetik. A probléma ott van, hogy a
darwini evolúciós elmélet szerint a fejlődési vonalon nem
tulajdonságoknak kellene szerepelniük, hanem kihalt fajoknak. Olyan
szervezeteknek, amelyek a felsorolt tulajdonságoknak csak egy
részével rendelkeztek, és nem a teljes kombinációval, aminek
következtében a természetes szelekció áldozataivá váltak. De ilyen
sikertelen próbálkozásokat megtestesítő, „átmeneti jellegű” fajokat
nem ismerünk, csak a sikeres végeredményt. Röviden azt is mondhatnánk,
hogy a molekuláris taxonómiai vizsgálatok nem erősítették meg
egyértelműen a darwini hipotézist.
Folyamatos vagy szakaszos fejlődés?
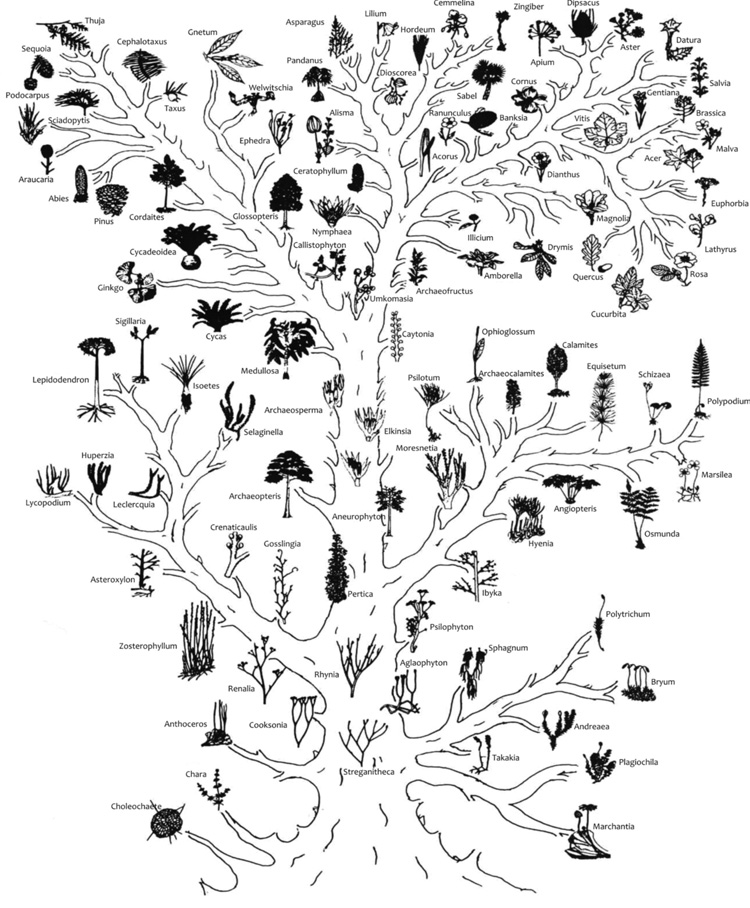
Ha már most a fésű alakú kladogramokat valódi fa alakú törzsfává
alakítjuk át
(1. ábra),
ahogyan azt Podani János tankönyvében látjuk (Podani, 2003),
rendkívül impozáns képet kapunk arról, hogy a Devon kor óta a
szárazföldi növények különböző csoportjai milyen sorrendben és milyen
származási viszonyban jelentek meg az evolúció során. Ez a törzsfa
szükségszerűen a mért és vizsgált genetikai távolságok léptékében
készült, és a fejlődést a darwini elmélet lépcsőzetes folyamatában
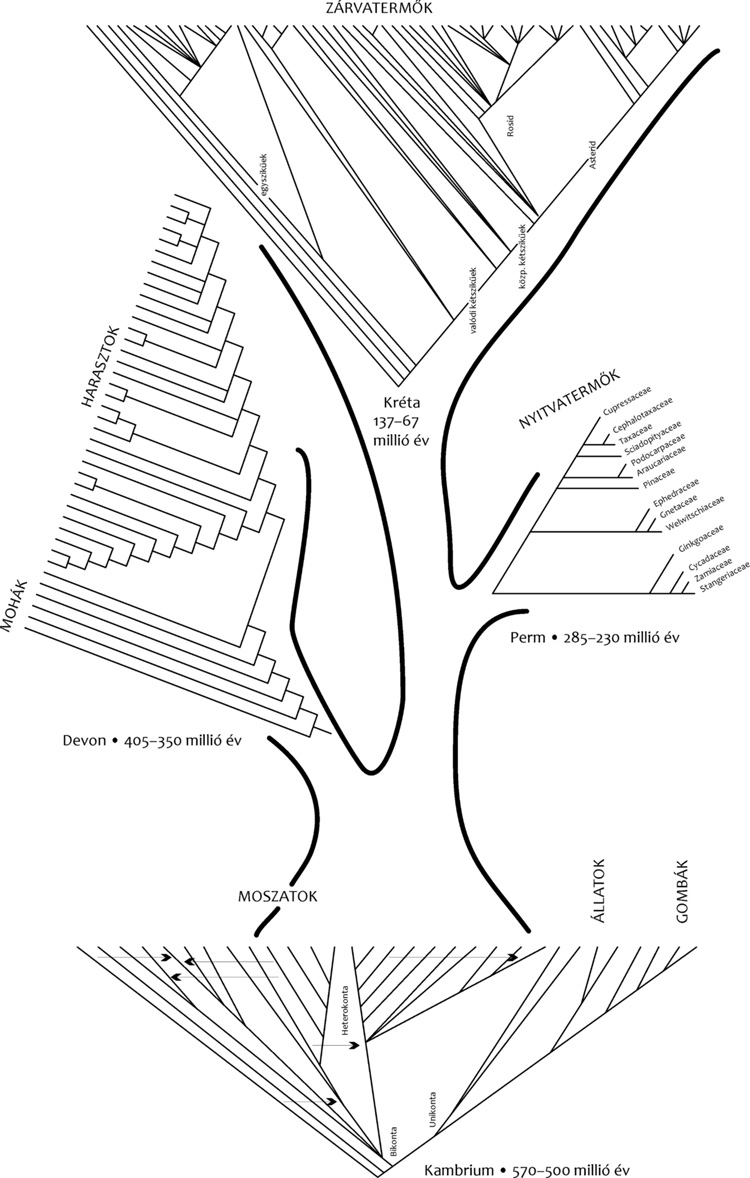
mutatja meg. Ha azonban ugyanezt a törzsfát a növénycsoportok
megjelenésének időarányos léptékében helyezzük el, ahogyan
megjelenésük a paleontológiai ismereteink szerint történt, merőben más
képet kapunk, amely arról szól, hogy a növényvilág evolúciója négy
nagy − időben elkülönülő − robbanásszerű szakaszra
tagolódik
(2. ábra),
amelynek
jellemzője, hogy mindegyik fejlődési szakasz kezdeti időszakát az új
alakok viszonylag gyors és gazdag megjelenése jellemzi.
A négy nagy evolúciós robbanás: 1.) az őstenger élővilágának
kialakulása, 2.) a növények partraszállása, 3.) a
nyitvatermő növények, 4.) a zárvatermők robbanásszerű megjelenése.
Az első robbanás: az őstenger élővilága
A négy robbanás közül az első a kambriumkori őstenger élővilágának a
kialakulása. Ez a korszak mintegy kétmilliárd esztendővel ezelőtt
kezdődött, de magát az őstenger élővilágát csak a mintegy félmilliárd
évvel ezelőtt keletkezett geológiai rétegekből ismerjük, mivel a
korábban keletkezett földtani rétegek a földkéreg nyomása alatt
átkristályosodtak, és a bennük tárolódott maradványok elpusztultak: a
„kozmikus titkosszolgálat” a korábbi dokumentumokat kevés kivétellel
megsemmisítette. A földi légkör ekkor még alig tartalmazott oxigént,
ezért hiányzott a ma létező ózonpajzs, amely az élő sejteket
megvédhette volna az UV-sugarak gyilkos hatásától. Ezért a
növényvilág egy sugárzásmentes bunkerbe kényszerült, ahol a bunker
szerepét a tenger látta el.
Az egysejtű élőlények fejlődésében mindjárt az elején két nagy csoport
elkülönülése figyelhető meg, az egyostoros és a kétostoros
egysejtűeké. Mindkét csoport tagjai úgy szerezték meg a fejlődésükhöz
szükséges sejtszerveiket és a fotoszintézishez szükséges
színtesteket, hogy más egysejtű élőlényeket faltak fel, melyek
sejtmagja, illetve szintestei tovább éltek és működtek immár a
gazdaszervezet belsejében mint azok saját szervei. Ez a jelenség az
ún. endoszimbiózis, amely számos alkalommal zajlott le, és különböző
egysejtű élőlények kialakulásához vezetett. A kétostoros sejtek nem
tudták megemészteni a felfalt színtesteket, amelyek még régi
sejtfalukat is megőrizték, és tovább folytatták fotoszintetizáló
tevékenységüket. Ezekből lettek a különböző algacsoportok, vagyis a
növények. Az egyostoros sejtek erősebb „gyomorsavval”
rendelkeztek, és sikerült megemészteniük a színtesteket is. Belőlük
fejlődtek a gombák és az állatok.
Az UV-sugarak eleinte mélyen behatoltak a tenger felszíne alá. Ezért
az első prokarióta és eukarióta növényi szervezetek 70−150 m
mélységben alakultak ki, ahová csak a sötétzöld fénysugarak jutnak
le, és ezért az első vízi növények a vörös színtestekkel rendelkező
cianobaktériumok és vörös algák törzséből kerültek ki. Később a
légkör magasabb oxigéntartalma már jobban megszűrte az UV-sugarakat,
és kialakulhattak azok az algatörzsek, amelyek 20−70 m mélységben
képesek fotoszintetizálni, ahol a fény kék színű. Itt a barna és sárga
színtestekkel rendelkező, ún. chromista algák váltak uralkodóvá, mint
a kovaalgák vagy a barnamoszatok. Még később jelent meg a
zöldmoszatok túlnyomó része, amelyek csak 20 m mélységig tudnak
fotoszintetizálni, és amelyek különösen a sekély tengerekben és a
partközeli zónákban jutottak vezető szerephez. Az egyes
algatörzseknek ez a mélységi rétegződése a tengerekben ma is
megfigyelhető. Végül megjelentek a csillárkamoszatok, amelyek a
szárazföldi létforma felé az átmenetet mutatják, mert ma is a
folyótorkolatok brakkvizében és az édesvizekben fordulnak elő.
Az őstenger élővilágának kialakulását azért kell robbanásnak
tekintenünk, mert a kambriumi őstengerben már szinte készen áll a
teljes evolúció, hiszen megtaláljuk a kilenc algatörzs mellett a
gerincesek kivételével mind a tizenhét állattörzset is. Sőt, az
evolúciónak abból a nyolc nagy lépéséből, amelyeket Maynard-Smith és
Szathmáry Eörs mutatott ki és elemzett (Maynard-Smith – Szathmáry,
1995), hat nagy lépés már az őstengerben megtörtént, gyakorlatilag a
szemünk elől elrejtve, hiszen ezekről fosszilis maradványok szinte
egyáltalán nem maradtak fenn. Ezzel kapcsolatos érdekes élményem
volt, hogy Dudich Endre professzor urat, − aki a zoológia kiemelkedően
legnagyobb szaktekintélye volt a 20. századi magyar biológiában −
kérdeztem, hogy ő hogyan látja az evolúciót. Ez nagyon bizalmas
kérdés volt, mert politikai szankciókat válthatott ki az 50-es években
Magyarországon. Ő azonban nem küldött el azzal, hogy fiatalember, maga
engem ne provokáljon, hanem őszintén válaszolta: „Nézze, én ebben
azért nem gondolkodom, mert korábban több éven át tengerbiológiával
foglalkoztam, többek közt a Nápolyi Tengerkutató Állomáson, és úgy
látom, hogy szinte a teljes állatvilág készen megtalálható a
kambrium kori őstengerben, és a gerinceseknek csak azok az osztályai
hiányzanak, amelyek a szárazföldi lét során jelentek meg.” Felfogása
cseppet sem egyedülálló. Simon Conway Morris Cambridge-i
paleontológus, a Royal Society tagja is a kambriumi robbanásról ír:
„Ma ugyan megfigyelhetők bizonyos átmeneti formák egyes fajok között,
és ebből arra lehet következtetni, hogy a múltban is léteztek ilyenek.
Mindazonáltal a végeredmény nagyon távol van a formák egy hézagmentes
mintázatától, ami lehetővé tenné, hogy egy nyomozó kiolvassa belőle
az élet „családfáját” egyszerűen úgy, hogy megkeresi a közbenső – élő
és kihalt – formákat, amelyek elvben összekötik a fajokat.
Ellenkezőleg, a biológusok a szerves forma diszkrét voltával és a
közbenső formák általános hiányával szembesülnek” (Conway Morris,
1998, 4.).
Ugyanezt erősíti meg James Valentine A rendszertani törzsek
eredetéről című tanulmányában: „Az élet fájának sok ága – nagy és
kicsi egyaránt – kriptogén eredetű, azaz rejtett eredetű, ősei
ismeretlenek. E hézagoknak egy részét bizonyosan az okozza, hogy a
fosszilis leletanyag nem teljes. De ez nem lehet az egyetlen
magyarázat egyes családoknak, a gerinctelenek sok rendjének, a
gerinctelenek összes osztályának és a metazoa összes törzsének
kriptogén természetére”. (Valentine, 2004, 35.)
Második robbanás: a szárazföld meghódítása
A második robbanás a devon korszakban következik be. Az őstenger
algáinak kétmilliárd éves fotoszintetikus tevékenysége kellett ahhoz,
hogy a földi légkör oxigéntartalma 10 %-ra növekedjék, ami már elég
vastag ózonpajzs kifejlődésére volt elegendő ahhoz, hogy a
szárazföldön is megvédje az élőlényeket az UV-sugárzástól. Nem
tudjuk, hogy a növények hogyan érzékelték, hogy már ki lehet jönni a
szárazföldre. Alig tudunk sikertelen kísérletről annak ellenére, hogy
a szárazföldi élet első kísérletéről van szó! Különösen szembetűnő, ha
ezt összehasonlítjuk a sikeres növények bámulatos sokfélesé-gének
robbanásszerű megjelenésével. Ne feledjük, hogy mindjárt kezdetben
olyan növényekkel találkozunk, amelyek már rendelkeznek kifejlett
bőrszövettel, rögzítő szervvel és hajtásrendszerrel, pedig a vízi
növénynek, ha szárazföldi életet kezd, meg kell alapoznia a növényi
test stabil egyensúlyi helyzetét a légtérben. Ehhez a hajtás és
ágrendszerek új szimmetria-viszonyait, új testformákat kell
kifejlesztenie, és mindezt a genetikai kódba beillesztve átörökíteni.
A szimmetriatípusok azonnal nagy változatosságban jelentek meg; szinte
egyszerre alakult ki a villás, az örvös, és a főtengelyes elágazási
rendszer, amelyek a devon kor óta eltelt több százmillió év alatt sem
változtak jelentős mértékben. A kialakult szerv- és szövetrendszerek
egyúttal megkapták a feladatukat is. A rögzítő szervnek meg kellett
oldania az anyagfelvételt, a szilárdítást végző hajtásrendszernek a
tápanyagok szállítását, az ágrendszernek pedig a légtérbe kifeszülve
meg kellett oldania az energia-felvételt és a gázcserét. A szexuális
folyamatok terén is jelentős előrelépés történt azáltal, hogy egyre
elterjedtebbé vált a heterospória, a kétivarúság, amely az öröklődést
genetikailag stabilabbá, a változékonyságot és alkalmazkodó
képességet pedig erősebbé tette. A heterospória azonban megkövetelte,
hogy a megtermékenyítés szárazföldi körülmények között is
üzembiztosan menjen végbe. A fejlődésnek ebben a stádiumában ez úgy
oldódott meg, hogy a hím és a női ivarszerv igen közel került
egymáshoz, és egyetlen harmatcseppben is megtörténhetett a
megtermékenyítés. A módszer sikerét mutatja az a hatalmas
alakgazdagság, amelyet korábban a Karbon kor sajátjának tekintettünk,
ám valójában már a Devon kor ötvenmillió éve alatt kialakult.
Hangsúlyoznunk kell, hogy bár a Devon korszak és az azt megelőző korok
(Ordovicium, Szilur) rétegeit meglehetős alapossággal vizsgálták át,
és vizsgálják ma is, keresve az első edényes növényi élet
bizonyítékait, „csalódottan kell megállapítanunk, hogy a legrégibb
leletek sem mondanak nekünk semmit az algaősökkel vagy bármely
mohacsoporttal való közös őssel fennálló kapcsolatról” (Podani, 2003,
31.) A növényi evolúció első és második nagy lépcsője között tehát
hiányzanak az átmenetet, az összekötő kapcsot jelentő élőlények,
vagyis a darwini fejlődéselmélet bizonyítékai.
Ezzel szemben a fejlődés robbanásszerű jellegét kitűnően szemlélteti a
kor geológiai rétegeiből előkerült fosszíliák óriási változatossága:
az ősi Rhynia flórát követő Zosterophyllumok, a valódi korpafüvek és
azok fává növő, erdőalkotó fajai a pecsétfák és pikkelyfák,
aljnövényzetükben a csipkeharasztokkal. Már a villás elágazású
növények uralmával egy időben megindul a száras növények
evolúciójának legjelentősebb vonala, a főtengelyes elágazású zsurlók
és páfrányok kialakulása. A molekuláris kutatások egyik legmeglepőbb
eredménye az volt, hogy az alakilag igen különböző zsurlókat és
páfrányokat egy leszármazási csoportba vonta össze.
Még a Devon vívmányai közé tartozik egy óriási fontosságú evolúciós
lépés: a mag kialakulása az előnyitvatermők képviselőjével (Archaeopteris).
A hatalmas alakgazdagság kifejlődésének hátterében jelentős
környezeti változások is lezajlottak, amelyek kedveztek a szárazföldi
növényvilág robbanásszerű kialakulásának, amilyen az egységes
őskontinens, a Pangaea kialakulása. Valamennyi szárazulat egyetlen
kontinenssé tömörült össze, és ennek a hatalmas szárazulatnak a
túlnyomó része hosszú évtízmilliókra a trópusi övezetbe került. Ebben
a trópusi környezetben alakult ki a páfrányok törzsének szinte
valamennyi csoportja a páfrányfáktól a vizek felszínét beborító
vízipáfrányokig.
Stephen Jay Gould paleontológus írja erről Az evolúció egyenetlen
sebessége című írásában (Gould, 1977) hogy „A legtöbb fosszilis faj
történetének két olyan jellegzetessége van, amely összeférhetetlen
azzal az elmélettel, miszerint fokozatosan alakultak ki: 1.) Hirtelen
megjelenés: valamely területen egy faj nem fokozatosan alakul ki
őseinek fokozatos átalakulása révén, hanem hirtelen jelenik meg,
méghozzá ’teljesen kialakult formában’. 2.) Stagnálás: a legtöbb faj
nem mutat határozott irányú változást földi létezése során. Amikor
megjelennek a fosszilis leletek között, ugyanúgy néznek ki, mint
amikor eltűnnek.” Gould itt nem azt állítja, hogy a fajkeletkezés nem
lehet kis lépésekből álló folyamat, például hibridizáció vagy
poliploidia útján, úgy, ahogyan azt Vida Gábor páfrányokon végzett
kutatásaival is bizonyította, hanem arra utal, hogy a fejlődés
nagyobb lépései, amit a fosszilis maradványok szintjén, és különösen
új típusok megjelenésekor tapasztalunk, nem a darwini evolúció szerint
történtek.
A harmadik robbanás: a nyitvatermők
Az evolúcióban a hirtelen nagy ugrás nem egyszeri, véletlen esemény
volt, amit a harmadik robbanás, a nyitvatermők kialakulása bizonyít.
Ennek előkészületei már a devon végén és a karbonban megtörténtek az
előnyitvatermők és a magvaspáfrányok kialakulásával. A mag
kialakulására számos kísérlet történik. A harasztok mindegyik
csoportjában megjelenik a mag, de csak a magvaspáfrányok vonalán
sikerül továbbvinni az evolúcióban. Ez nem csoda, hisz az életképes
mag kifejlődéséhez hat fontos kritériumnak kellett együttesen
teljesülnie. 1.) A különnemű spóráknak, vagyis a heterospóriának. 2.)
A női spóratartóban a spórák számának egyre csökkennie kell. 3.) A
következő lépés, hogy a női ivaros nem-zedék benne maradjon a női
spóratartóban, és ez az ivartalan életfázisú szervezeten fejlődjön ki.
4.) Ezután a női spórának új védelmi szerkezetet kellett kialakítani
azáltal, hogy egy burokkal vette körül magát (integumentum) és 5.)
ennek csúcsán kialakítania a virágporszemet felfogó készüléket (mikropüle).
6.) Ezzel párhuzamosan a következő hat fejlődési folyamat megy végbe
a hím ivarszerben: a.) a hímivarsejt elveszti ostorait, b.) benne
marad a mikrosporangiumban, amely c.) ellenálló fehérjeburokkal
veszi körül magát, és d.) alkalmas a széllel való szállításra, továbbá
e.) a mikropüle beléptető kódjára helyes választ adva f.)
pollentömlőt fejleszt, és így juttatja el a hímivarsejteket a
petesejthez.
A darwini elmélet szerint ez az utódlási rendszer a változékonyság
véletlenszerű tapogatózása során, évmilliók alatt jött volna létre.
Nincs kielégítő válaszunk arra, hogy miért a karbon időszakban megy
végbe mindez, amikor még nincs rá szükség? Honnan „tudták” a
növények, − hiszen valamennyi csoport megkísérelte a mag
kifejlesztését −, hogy a Perm időszakban ez lesz a túlélés eszköze? A
Karbon időszak trópusi „jóléte” után ugyanis a Permben váratlanul
beköszönt az éghajlat szárazodása. A Pangaea kisodródik a nedves
trópusokról a száraz szubtrópusi öv alá, miközben megkezdődik a
kontinens feldarabolódása. Ennek kényszerítő hatására a
szárazságtűréshez való alkalmazkodási módok és formák különböző
területeken, egymástól elszigetelődve, eltérő módon jönnek létre. Meg
kell oldani a szárazságtűrés különböző formáit, például a szállító
szövetrendszer tökéletesedését, a párologtató felület csökkentését,
és ekkor válik igazán a siker kulcsává a mag, amely képessé teszi az
utódot a kedvezőtlen időszak átvészelésére, és a magvas növényeket
arra, hogy átvegyék az uralmat a szárazföldi térségeken. A
megtermékenyítés korábbi − vízcseppben való − módja a száraz éghajlat
alatt bizonytalanná válik, a mag kialakulását létrehozó stratégia, a
széllel szállított − „léghajóban” utazó − hímivarsejtek, a
szélmegporzás lesz a megtermékenyítés uralkodó formájává.
A kontinensek szétsodródása következtében a nyit-vatermők három
különböző csoportja már a fejlődés korai stádiumában elvált egymástól,
és ezt a molekuláris vizsgálatok alapján készült kladogramok is
megerősítették. Ezek: a pálmaszerű testfelépítést mutató
cikász-bennetitesz vonal, amelynek széles levelein fejlődnek ki az
ivarszervek; a CordaitesGinkgo-fenyők vonal, csúcsán a toboztermő,
tű vagy pikkelylevelű fenyőkkel, amelyeken az ivarszervek a
tengelyképleteken helyezkednek el; és végül a Gnetum-féléknek három
igen különböző egységből álló csoportja. A nyitvatermők nagy része
mára kihalt, és a fenyőféléken kívül valamennyi ma élő csoport a
túlélés utolsó stádiumában, a kipusztulás határán levő élő kövület. A
Perm–Triász–Júra időszakok nyitvatermő flórájának nagy
változatosságát mutatják a kihalt csoportok is. A száraz klíma nem
kedvez a fosszilizációnak, és ez meg is mutatkozik a sokkal kisebb
számú nyitvatermő maradványon. Ezek azonban ahelyett, hogy „betömnék”
a ma élők között tátongó réseket, tovább fokozzák a változatosságot.
A kihalt szárazságtűrő kislevelű magvaspáfrányok (Glossopteris,
Coristosperma, Pentoxylon Caytonia), és a pálmaalakú
Bennetites igen eltérő formái mind egy-egy külön fejlődési vonalat
látszanak képviselni. Ennek eredménye, hogy a nemrég még egyetlen
rendszertani törzsnek tekintett nyitvatermőket ma legkevesebb öt
törzsre bontva tárgyaljuk, ami azt jelenti, hogy hiányzanak közülük az
összekötő elemek.
A paleontológusok már Darwin életében azt kifogásolták, hogy a
fejlődés nagy lépcsői közül hiányzanak azok a kis lépcsők, amelyeknek
a fokozatos fejlődést kellene igazolniuk. Darwin azzal védekezett,
hogy a geológiai rétegek feltáratlansága miatt kevés fosszíliát
ismerünk, de a földkéreg feltárása során várhatóan elő fognak kerülni
azok a maradványok, amelyek a fejlődés még ismeretlen hiányzó
láncszemei. Ez a hiányzó láncszem, a missing link volt a varázsszó,
amellyel a darwini elmélet problémáinak megoldását elnapolták. Darwin
maga is érezte, hogy elméletének ez a gyenge pontja, mert azt írja:
„Korábban rengeteg közbenső változatnak kellett léteznie a Földön.
Akkor miért nincs tele minden geológiai formáció és réteg ilyen
közbenső láncszemekkel? Bizonyos, hogy a geológia nem tárt fel
semmilyen fokozatos szerves láncot. Talán ez a legnyilvánvalóbb és
legsúlyosabb kifogás, ami felhozható elméletem ellen.” (Darwin, 1996,
227.) De már 1952-ben Nils Heribert-Nilson azt írta (1952, 1212.): „A
paleobiológiai tények alapján még karikatúrát sem készíthetünk az
evolúcióról. A kövületi anyag most annyira egységes, hogy az átmeneti
alakok láncszemeinek hiányát nem lehet többé szegényesen rendelkezésre
álló kövületi anyagokkal magyarázni. A hiányok valóságosak; soha sem
fognak kiegészülni”. Véleményének helyességét igazolta az idő, mert
huszonöt évvel későbben David Raup, a chicagói Field Múzeum
paleontológusa hasonlóan ír (Raup, 1979, 25.) „Darwin óta körülbelül
százötven év telt el, és a fosszilis leletanyag jelentősen bővült. Ma
negyedmillió fosszilis fajt ismerünk, de a helyzet nem sokat
változott. Az evolúcióra utaló leletanyag még mindig meglepően
egyenetlen. A sors iróniája, hogy ma kevesebb példánk van az evolúciós
átmenetekre, mint Darwin idejében volt.” De még ennél is tovább megy
Nils Eldredge, az American Natural History Museum paleontológusa: „Mi
paleontológusok azt mondtuk, hogy az élet története alátámasztja a
fokozatos adaptív változás elméletét, miközben egész idő alatt tudtuk,
hogy ez nem igaz.” (Eldredge, l985, 144–145.)
Ez olyan megdöbbentő beismerés, amihez fogható nem olvasható minden
nap a tudományos irodalomban.
A negyedik robbanás: virágba borul a Föld
És végül jön a növényvilág fejlődésének utolsó nagy robbanása: a
zárvatermő növények világuralmának kialakulása. Náluk tovább
tökéletesedik a szállító szövetrendszer, az utódvédelem a zárt
magházzal, és az embriónak különleges, „roboráló” tápszövettel való
ellátása, a megtermékenyítés pedig a színpompás és csodálatosan
formagazdag virágtakaró kialakulásával és az állatmegporzással. Ezek
óriási horderejű változások, amelyekkel a termőhelyek benépesítéséért
és a forrásfelosztásért folyó versengésben a zárvatermők valósággal
„állva hagyják” a nyitvatermő növényeket, amelyek kiszorulnak a hideg
éghajlatú zónákba és a tápanyagban szegény talajokra.
Első lépésként meg kell keresnünk e színpompás gazdagság eredetét. Már
a nyitvatermők őseinek keresése is zavarba ejtő nehézségekkel volt
terhes. Még ennél is nehezebb a zárvatermők őseit megtalálni.
Származásukat már Darwin is „irtóztatóan titokzatosnak” ítélte. A
nagy reménnyel várt molekuláris kutatások eredményei inkább fokozták
a bizonytalanságot. Kimutatták ugyanis, hogy a potenciális ősként
leginkább számon tartott nyitvatermők, a Gnetum-félék,
genetikailag a fenyőkhöz állnak közel, és semmiképpen nem lehetnek a
zárvatermők ősei. Sokáig a cikász-félék rokonságában keresték a
zárvatermők virágának ősét (euanthium-elmélet), újabban azonban a
Coristosperma-típusú kihalt magvaspáfrányok csoportjának
valamelyik – eddig ismeretlen – tagjában remélik a zárvatermők ősét
megtalálni.
Nem sokkal jobb a helyzet, ha a zárvatermők ősi, kiinduló csoportjait
akarjuk meghatározni.
Mivel a mai nyitvatermők körében a fás szár kizárólagos, az a
legelfogadottabb feltételezés, hogy az ősi zárvatermők is fás növények
voltak. Így esett korábban a választás a liliomfákra és primitív
rokonaikra (Winteraceae, Degeneriaceae), de ezek kiinduló
pozícióját a genetikai vizsgálatok nem igazolták. A molekuláris
kladisztikai vizsgálatok szerint a legősibb típus az Amborella
nevű cserje. Vele két gondunk van. Az egyik az, hogy az Amborella
genomjának ősi jellegét részben a benne található májmohagéneknek
köszönheti, amelyek adventív gének is lehetnek, amit valószínűsít,
hogy a májmohák a fejlődés oldalágán vannak. A másik gond az
Amborella elszigeteltsége és reliktum termőhelye. Valószínűtlen,
hogy ilyen pozícióból viszonylag rövid idő alatt világuralmi
pozícióba kerülhetett a növényvilág legnépesebb csoportja. A
zárvatermők származási vonalának második helyén a tündérrózsa-féléket
találjuk, amelyek mind lágyszárú vízinövények. Esélyüket növeli, hogy
a nyitvatermők között nincsenek, a páfrányok között alig vannak
lágyszárú vízinövények, vagyis monopolhelyzetben voltak a mélyebb
víztestek benépesítésében. Ha ökológiai ismereteink birtokában
megkíséreljük elgondolni, hogy hol lehetett esélyük a zárvatermőknek
egy fenyvesekkel és ginkgoerdőkkel borított tájba behatolni, akkor a
folyómedreket és az állóvizeket jelölhetjük meg elsőként ilyen
termőhelyeknek.
Az ősi zárvatermők dilemmája nemcsak az, hogy nem találjuk a „közös
őst”, hanem hogy ezek, − mint láttuk − mind morfológiailag, mind
származástanilag igen távol állnak egymástól. Egy rózsa és egy
napraforgó genetikailag és szervezetileg is közelebb áll egymáshoz,
mint egy tündérrózsa az Amborellához. Még az a gondolat is
felmerült nemrégiben, hogy van-e közös ősük a zárvatermőknek
egyáltalán, vagy az ősi zárvatermők különböző ősök leszármazottai-e?
Mindeme kérdéseknek egyetlen biztos következtetésük van, éspedig az,
hogy a zárvatermők evolúciójának alaplépései is ugrásszerűek voltak,
és nem a darwini modell szerint történhettek.
|
|
|
Egy új információs kapcsolat, az
együttes fejlődés
Különösen fontos új mozzanat a növény- és állatvilág korábbi egyoldalú
kapcsolatrendszerének kölcsönössé válása. A szárazföldi lét első
kétszázmillió éve során a növényvilág élelem, fészkelő- és búvóhely
formájában egyoldalúan szolgálta ki az állatvilágot. A növény- és
állatvilág szaporodásbiológiája gyakorlatilag független volt
egymástól. Az első őscserjék kialakulásától kezdve az első
zárvatermők megjelenéséig a növényi petesejt megtermékenyítése vagy
vízcsepp segítségével, vagy szél általi közvetítéssel ment végbe. A
zárvatermők az első nagy növénycsoport, amelynek szaporodásában az
állatvilág különböző csoportjai meghatározó szerepet játszanak. Ezzel
a fontos lépéssel következik be a növényvilág szexuális értelemben
vett felnőtté válása − ha úgy tetszik: emancipációja, − amikor
kiszabadul a „szél úrfival” kötött kényszerű házasság százötvenmillió
éves monotóniájából, és az utolsó száznegyvenmillió évet már
egyenrangú partnerkapcsolatban éli meg az állatvilággal, amely
változatos megjelenésű „kérők” hadával jelentkezik a megporzás és
megtermékenyítés lebonyolítására. A zárvatermők válasza kettős: a
növények kiöltöznek, és választanak. A kialakuló színes virágtakaró
szerepe ugyanaz, mint a menyasszonyi ruháé: a sikeres nász
elősegítése. A nagyszámú és sokféle megjelenésű „kérő” igen rövid idő
alatt kialakította a „ruhaszalonok” versengését. Az egyszerűbb fazon
vonzását illatszerrel pótolták, hiszen az udvarlók egy része −
összetett szeme ellenére − rövidlátó, viszont kifinomult szaglással
rendelkezik. A virágok versengése más vonatkozásokra is kiterjed. A
zigomorf virág megjelenése praktikus célokat szolgál: leszállópályát
kínál a látogatónak. Az orchideák körében a megporzási aktus az
intimitásig finomulhat. Az Oncidium és Catasetum nevű
orchideafajok virágzása egybeesik bizonyos trópusi darázsfajok
nászrepülésével, és a virágok a párzó nőstény illatával
„kedveskednek” a hímeknek. A bangók a párzó nőstény képét öltik
magukra. A Coryanthes-kosbor bódító illatú fürdővel várja a
látogatókat. És ezt az egész folyamatot, amely a növény- és rovarvilág
szexuális kérdés-felelet játéka, nevezzük együttes fejlődésnek, azaz
koevolúciónak.
A koevolúcióval kapcsolatban több olyan kérdés merül fel, amelyek
megválaszolásával adósok vagyunk, vagy legalábbis a válaszaink nem
kifogástalanok. A növényvilág mindjárt a kezdeteknél két nagyon
fontos fejlesztéssel lép be ebbe az együttmű-ködésbe. Az egyik a
virágtakaró szín- és alakgazdagságának robbanásszerű változatossága.
Az első kilenc zárvatermő rendben tucatnyi virágtípus jelenik meg,
egészen a forrt virágtakarójú csapdavirágokig. A másik a csalogató
jellegű kémiai anyagok korai megjelenése: a vízinövényeken kívül
valamennyi primitív zárvatermő növény illóolajokat termel. Egyelőre
nincs kielégítő magyarázatunk arra, hogy mi módon történt meg az
információátadás, illetve információáramlás az egymásra utalódó
növény- és állatcsoportok között. Újra jelentkezik az evolúció állandó
dilemmája a kezdetről: a tyúk-tojás dilemma, vagyis, hogy ki kezdte az
alkalmazkodást? A növény idomította-e a virágját a rovar
szájszervéhez, vagy fordítva, a rovar szájszerve idomult a már
kialakult virágszerkezethez? Például az Angraecum sesquipedale
nevű orchidea 32 cm hosszú sarkantyújában levő nektárhoz egy olyan
lepkefaj fér hozzá, amelynek pödörnyelve kinyújtott állapotban
hatszo-rosa az állat testhosszának. Érdekes lenne kiszámítani, hogy
hány véletlenszerű kísérletre volt szükség ilyen különleges testarány
kialakításához. Ez a kapcsolat ellentmondani látszik az amerikai
zoológiában a 70-es évektől elterjedt nézetnek, hogy a növények
virágai alkalmazkodtak volna az állatok szájszerveihez, amire a fiatal
szigetflórákban, például Hawaii flórájában véltek példákat találni.
Arra sincs magyarázatunk, hogy miért kellett a zárvatermő virágnak
nemcsak hasznosnak, hanem szépnek, sőt, egyre szebbnek lennie. Ha
engem valami az evolúcióban arra inspirál, hogy egy Teremtőben
gondolkozzam, az a természet rendkívül magas szintű esztétikája, amely
elsősorban a növények virágaiban és virágzataiban nyilvánul meg.
Lehet, hogy ez nincs megtervezve, mégis úgy néz ki, mintha meg lenne,
annál is inkább, mert hiszen megvannak azok az alkalmazkodott
állatcsoportok, amelyek ezt keresik. De vajon van-e a rovaroknak
esztétikai érzékük? Úgy tudjuk, hogy nincsen. De akkor mire való ez a
tékozló gazdagság? Az embernek az a határozott benyomása, hogy az
evolúció megalkotó-jának öröme telt az alkotásban.
A koevolúció másik fontos kérdése az információáramlás. A zárvatermők
megjelenéséig a növény és az állatvilág evolúciója párhuzamosan,
nagyjából függetlenül, külön-külön zajlott. Most azonban a két
evolúciós folyamat informatikailag összekapcsolódik, de nemcsak
összekapcsolódik, hanem rendkívüli mértékben fel is gyorsul, annak
ellenére, hogy a két folyamat összehangolódása − véletlen keletkezést
feltételezve − feltétlenül több időt igényelne. Ezzel szemben az alsó
krétától a felső krétáig eltelt negyvenmillió év alatt kialakul
csaknem valamennyi ma élő növénycsalád, ami több mint háromszáz
növénycsaládot jelent. Természetesen nem mindegyik a mai
fajgazdagságával van képviselve, de mind-egyiknek az első képviselői
már megvannak. És közben mindegyik család tagjai megtalálják a
beporzó ágenseiket, amelyek részére testre szabott tápanyagot,
különböző cukorösszetételű nektárt, zsír- és fehérjetartalmú
tápszövetet, különböző illóolajokat tartalmazó csalogató anyagot
„készítenek”. Ezek között bonyolult felépítésű vegyületek vannak,
amelyeket az anyagcsere melléktermékeiként kell előállítani, amiről
tudjuk, hogy nem megy egyik napról a másikra. Ehhez még vegyük hozzá
azt a körülményt, hogy véletlenszerű keletkezés esetén fennáll a
veszély, hogy hibás anyag keletkezik, amit „a rosszul kemizáló
növénnyel együtt ki kell dobni”. Szegény növény már régen kipusztult,
mire a megfelelő vegyületet „kikísérletezte” volna. Ha megkíséreljük
kiszámítani, hogy ennek a koevolúciós folyamatnak mi a valószínűsége
véletlenszerű keletkezés esetén, akkor rendkívül kicsiny számot
kapunk. K-számították, hogy a sikeres kísérletek előállítására nem
negyvenmillió, de még négyszázmillió év − vagyis a teljes szárazföldi
élet tartama − sem lenne elegendő.
Az evolúció forrása és iránya
Mivel az evolúció egy folyamat, elkerülhetetlen annak a kérdésnek a
feltevése, hogy mi a hajtóereje ennek a folyamatnak? Ehhez az élő
anyag változékonysága egyedül kevés. Kell hozzá a meghódítható
környezet, amely legalább minimális tápanyagkínálatot biztosít az élő
szervezetek számára. Ezt a tápanyagkínálatot a növények nemcsak
felhasználják, hanem tovább gazdagítják, vagyis tevékenységükkel
elősegítik az evolúció továbbhaladását. A növényzet nem vakon sodródik
az evolúcióval, ahogyan azt Dawkins szeretné elhitetni, hanem aktív
részese a fejlődés folyamatának. Kezdve ott, hogy a növények az
őstenger lakóiként megtermelték a földi légkör oxigénjét, majd
folytatva azzal, hogy a kontinensek holt kőzetein létrehozták a
termőtalajt, egy olyan képződményt, amely − ismereteink szerint −
egyedül a Földön található meg az egész Univerzumban. A növényvilág e
szerepvállalásai nélkül nincs evolúció!
A zárvatermők kialakulásával elég nagy élő mintaszámmal rendelkezünk
ahhoz, hogy megkockáztassuk a kérdést: van-e iránya a fejlődésnek?
Furcsa belegondolni abba, hogy az ember, aki maga is egy fejlődés
eredményének tekinti önmagát, nem találja ellentmondásnak, ha azt
hiszi, hogy ő találta ki a fejlesztést, amelynek természetesen van
iránya és célja. A zárvatermők fejlődési sorain végigtekintve azt
látjuk, hogy miközben a növény- és rovarvilág szexuális
összecsiszolódása folyik, ezzel egyidejűleg fokozatos erőfeszítések
történnek a növények részéről, hogy a megtermékenyítés folyamatát
technikailag és technológiailag is egyre biztonságosabbá, és
ugyanakkor egyre gazdaságosabbá is tegyék. Így lesz a genetikai
leszármazási sorban egymás után következő családokban a sok porzóból
kevés, a sok termőlevélből egy- vagy kétüregű magház, az embriózsák
kettős falából egy, a vastag nucelluszból vékony, a felsőállású
termőből alsóállású stb.
Példaként érdemes a fejlődés alapján álló tündérrózsának és a
virágfejlődés egyik csúcsát jelentő orchideáknak a virágát és
megtermékenyítési mechanizmusát összehasonlítani A tündérrózsa
virágát sok színes virágtakaró teszi látványossá, benne nagyon sok
porzót és termőlevelet találunk. A termőlevelekben sok magkezdemény,
amelyek együttes száma elérheti az egy-kétezret. A teljes
megtermékenyítéshez valamennyi termőlevélre legalább annyi
virágporszemnek kell kerülnie, amennyi magkezdemény van a magházban.
Ez szinte soha nem történik meg, a virágban mindig maradnak sterilen
maradt magkezdemények.
Az orchidea virágot mindössze hat színes virágtakaró borítja. A
virágban egyetlen porzó és egyetlen, két termőlevélből összenőtt
magház található. Ennek 180 fokos elfordulásával a legnagyobb
lepellevél ajakká módosul, és nektárt tartalmazó sarkantyúban
folytatódik, egyszerre kínálva leszállópályát és táplálékot. Az
egyetlen porzó ránő a bibére, és együttesen egy technológiai
remekművet, az ivaroszlopot hozzák létre, amelyben többféle
munkamegosztás van. A bibe egyik karéja ragacsos váladékot termel,
amellyel a porzó pollenzsákjában levő összes virágport egyetlen
ragadós pollentömeggé formálja. Ez egy nyéllel és a tövén levő
ragasztótesttel egy buzogány alakú képződményt hoz létre, az ún.
pollináriumot, amely így egyben kerül át a másik virágra. Repülés
közben a légáramlás szárító hatására a pollinárium nyele vizet veszít,
lehajlik, s ezzel pontosan a következő virág bibéjének magasságába
kerül, ahol mérnöki pontossággal meg is ragad, vagyis a megporzás a
teljes virágpormennyiséggel egyszerre megtörténik. Ekkor, és csak
ekkor indul meg a magházban a magkezdemények „legyártása”. Mialatt a
bibéről a pollentömlőfolyam leér a magházba, kialakulnak a
magkezdemények, amelyek száma négyezertől négymillióig terjedhet, és
megtörténik a magkezdemények tömeges megtermékenyítése. Bámulatos,
hogy ez a mechanizmus ugyanilyen pontossággal működik a trópusi
köderdők mohapárnáiban élő miniatűr orchideákon is, amelynek virága
1,5 mm átmérőjű, és mikroszkopikus méretű bogarak porozzák be. A
megtermékenyült toktermésben kifejlődő porszerű, apró magvak rendkívül
könnyűek, a szelekkel egészen a sztratoszférába is eljutva óriási
távolságokat képesek berepülni. A csíraképességüket sokáig megőrzik,
amire szükségük is van, mert „anyjuk útravaló nélkül bocsátotta el
őket”: az embriók egyetlen sejtnyi tápszövettel sem rendelkeznek!
Látszólag a legnagyobb gondosság a legnagyobb gondatlansággal párosul
az orchideák utódgondozásában. A felnőtt orchideanövények nagy része
gombafonalak segítségével veszi fel a vizet és a tápsókat, de ez nem
lehet elegendő információ ahhoz, hogy erre a lehetséges együttélésre
alapozva bocsássa el a növény a világba az utódait. Márpedig ez
történik. Az orchideanövény „számít arra”, hogy az útravaló nélkül
elbocsátott „bébi” meg fogja találni azokat a gombafonalakat, amelyek
„önkéntes dajkaként” majd felnevelik. Ez a „számítás” úgy van
bekódolva a magba, hogy csak gombafonal jelenlétében csírázik ki. Az
orchideák tehát kettős koevolúciós együttműködésre szerződtek, a
megporzásra az állatvilággal, a terjesztésre a gombavilággal. És ez
nem egyedüli eset. A hüvelyesek például mindkét funkcióra az
állatvilággal szerződtek, csak más-más állatcsoportokkal.
A zárvatermők fejlődésében felfedezhető további érdekes jelenség a
különböző helyeken kialakuló környezeti kihívásokra adott azonos vagy
hasonló adaptációs válaszok, párhuzamos fejlesztések a legkülönbözőbb
rokonsági körökben, amely jelenséget homopláziának nevezzük. Mintha a
különböző zárvatermő családok tarsolyában ugyanaz a haditerv vagy
használati utasítás lenne elrejtve a környezet azonos típusú
támadásaival szemben. Ez azért feltűnő, mert a zárvatermők kialakulása
közben igen nagyarányú felszíni átrendeződések történtek a Földön.
Mintegy nyolcvanmillió évvel ezelőtt kialakul az Atlanti-óceán
medencéje, szétsodródnak a kontinensek, és ezzel együtt a zárvatermők
fejlődése előbb négy, majd hat földrészen, jelentős mértékben
elszigetelődve, egymással párhuzamosan folytatódik. Ennek ellenére a
különböző kontinenseken a különböző genetikai és morfológiai
adottságokkal rendelkező növénycsaládok teljesen hasonló adaptációs
válaszokat adnak a környezeti kihívásokra. Legközönségesebb példája
ennek a szárazsághoz való alkalmazkodás: a törzs- és levélszukkulencia,
a pozsgás szövetek kifejlesztése. Ez azért is feltűnő, mert a
nyitvatermők nagy része is száraz éghajlat alatt élt, mégsem tudtak
víztartó szöveteket kifejlesz-teni. A Föld hat nagy sivatagi
övezetének meghódítására legalább húsz zárvatermő család fejlesztett
ki pozsgás növénycsoportokat, mindenütt másokat, megjelenésükben mégis
megdöbbentően hasonlókat.
De hasonló párhuzamos adaptációt látunk a rovarfogásra, illetve
fehérjeemésztésre való specializálódás vagy a magashegyvidéki
életformához (auxintermelés hőkorlátozása UV-sugárzás ellen) való
alkalmazkodás esetében is.
A fejlődés törvényszerűségei
Végigtekintve az evolúció során tapasztalt jelenségeken,
elérkezettnek láthatjuk az időt arra, hogy megfogalmazzuk az evolúció
néhány törvényszerűségét.
1.) Megállapítható, hogy a fejlődés szakaszos, és minden szakasz két
fázisból áll: egy revolúciós szakaszból, amelyet a hirtelen megjelenő
nagy formagazdagság jellemez, és egy evolúciós szakaszból, amely
hosszan tartó, és amelynek során a keletkezett típusok alkalmazkodnak
a környezeti feltéte.lekhez, térben elhelyezkednek és felosztják
egymás közt a rendelkezésre álló forrásokat, sőt, azokat tovább is
fejlesztik. Erre a második fázisra tekinthetjük érvényesnek a darwini
természetes szelekciót olyan értelemben, hogy azok a növények, amelyek
a forrásfelosztásban nem képesek az adottságaikat érvényesíteni, vagy
kipusztulnak, vagy elvándorolnak, vagy olyan genetikai változáson
mennek keresztül, például hibridizáció vagy poliplodizáció útján, ami
megnöveli a versenyképességüket. Úgy is mondhatnánk, hogy minden
szakasz egy kreatív és egy adaptív, vagy másképpen fogalmazva egy
rövidebb nem-darwini és egy hosszabb darwini fázisra tagolódik. A
revolúciós fázis genetikai történéseit több vonatkozásban nem ismerjük
kielégítően, az evolúciós fázis genetikájának számos technikáját jól
ismerjük, például Heribert-Nilson említett könyvéből, aki először
állított elő a természetben élő növényfajt másik két élő faj
keresztezésével. A 60–70-es években Vida Gábor derítette ki több
páfrányfaj hibrid eredetét, és sikeresen reszintetizálta a szülőkből
az utódfajt.
2.) Utolsó pár előre fuss! A fejlődés második törvényszerűségének ezt
a nevet adtam egy régi társasjátékról, amelyet biológusok,
geográfusok és geológusok korábban még gyakran játszottak az egyetemi
terepgyakorlatok estéin – amikor még jutott pénz terepgyakorlatokra az
egyetemi oktatásban. A törvényszerűség lényege az, hogy az új
szakasz növénycsoportjai nem az előző szakasz legfejlettebb, hanem a
legprimitívebb csoportjai közül fejlődnek ki. Ez az összefüggés mind
a nyitvatermők, mind a zárvatermők lehetséges őseinél is nyomon
követhető, és különösen világosan látszik a legfejlettebb
nyitvatermők, a Gnetum-félék esetében.
3.) A harmadik törvényszerűség, hogy a növényvilág aktív résztvevője a
fejlődésnek. A növények nem passzív elszenvedői a fejlődést kiváltó
környezeti változásoknak, hanem maguk is hozzájárulnak a fejlődést
előmozdító átalakulásokhoz, az új feltételek kialakításához. A
növényzet és környezet partnerkapcsolata lehet az oka annak, hogy a
növényvilágban kevés az evolúciós selejt. A véletlenszerű
kísérletezés szükségszerűen nagyon sok hulladékot eredményez. A
növényvilág evolúciójából hiányzik a nagy mennyiségű hulladék. Szinte
nincs selejt. De különben is: az evolúció teljes félreértése azt
gondolni, hogy a kihalt fajok sikertelen evolúciós kísérletek
voltak. Ezek a maguk idejében túlnyomórészt sikeres fejlődés
eredményei voltak, s évmilliókon keresztül kontinens nagyságú
területeken alkottak erdőségeket és halmozták fel például
szénkészleteink anyagát. Kipusztulásukat természeti katasztrófák,
vagy olyan gyors adaptációs kényszerek okozták, amelyekhez
specializálódott genetikai anyagaik már nem voltak elég rugalmasak,
vagy egyszerűen elfáradtak, elöregedtek, amire a ma élő flórában is
látunk példákat. De ezek nem evolúciós selejtek! Vagyis: a
véletlenszerűségnek egyszerűen hiányza-nak a bizonyítékai. És
valószínűleg azért, mert a növényvilág nemcsak alkalmazkodik a
környezethez, hanem át is alakítja a létfeltételeket. Előbb a maga
számára teszi alkalmassá, de ezen túlmenően olyan átalakításokhoz is
hozzájárul, amelyek a magáénál fejlettebb közösségek kialakulását
segítik elő. Ez a természetes szukcesszió, mely folyamat hasonló az
emberi társadalmak fejlődéséhez, ahol, ha a termelési eszközök
fejlettsége meghaladja a termelés színvonalát, akkor a társadalomban
gazdasági és technológiai váltás következik be.
Ez az összefüggés azért is fontos, mert a tudomány és a társadalmi
fejlődés között egy hasonló „koevolúció” létezik, mint a
természetben, vagyis a társadalom jelenségei visszahatnak a
tudományos gondolkodásra. Egy tudós tudományos gondolkodását
többnyire jelentősen befolyásolja az a társadalmi környezet, amelyben
él (vö. Borhidi, 2002). Darwin elméletének megszületésében is szerepet
kellett hogy játsszék az a társadalmi környezet, amelyben élt; az a
kiélezett és könyörtelen verseny, és a társadalmi igazságtalanságok
tömege, amelyek a korai kapitalizmus 19. század közepének Angliáját
jellemezték, s amelyeket nagy hűséggel ír le Charles Dickens a
regényeiben. Darwin elmélete többek közt azért lett oly gyorsan és
olyan széles körben elfogadott, mert ez az igazságtalan társadalom
hálás volt azért, hogy azt mondhatta: mi csak azt csináljuk, ami a
természetben folyik. A társadalom úgy érezte, hogy Darwin természetes
szelekciós elmélete felmentést ad a kollektív felelősség alól.
Darwinnak ez valószínűleg nem állt szándékában, de hogy elméletét a
társadalom, a köztudat és a politika erre is felhasználta,
vitathatatlan.
4.) Az evolúció negyedik törvényeként azt mondhatjuk ki, hogy a
fejlődésnek van iránya. Ezt ugyan Dawkins és a neodarwinisták
tagadják, de amikor élőlényeket fejlettség szempontjából
összehasonlítunk, a fejlődésről törzsfát készítünk, olyan szavakat
használunk, mint primitív és levezetett, akkor bizony irányultságról
beszélünk. A fejlődés irányát pedig egy szigorú, de nem vak fizikai
törvény szabja meg, az anyag és energia megmaradásának törvénye. Az
élővilág azzal az anyaggal és energiával gazdálkodik, amit kap. Az
egész evolúció arról szól, hogy az élővilág a saját lét- és
fajfenntartási funkcióit egyre kevesebb anyagból, egyre jobb
technológiai színvonalon, egyre biztonságosabban és egyre kevesebb
energia felhasználásával oldja meg. És ez igaz, akár a megporzás és
megtermékenyítés, akár a táplálkozás biológiájára gondolunk. Elég itt
utalnunk a szállítószövetek tökéletesedésére vagy a tündérrózsa és
orchidea korábban elmondott példájára.
Mi a fejlődéselmélet? Tudomány vagy ideológia?
Érvek és vélemények
Miután láttuk a saját érveinket, nézzük meg, hogy más tudósok hogyan
vélekednek a biológiáról és benne a darwini evolúciós elméletről:
Richard Dawkins (biológus): „A biológia olyan bonyolult dolgok
tanulmányozása, amelyek azt a benyomást keltik, mintha valamilyen
céllal megtervezték volna őket.” (Dawkins, 1986, 1.)
Francis Crick (Nobel-díjas biológus) Cambridge University: „A
biológusoknak nem szabad megfeledkezniük arról, hogy amit látnak, azt
senki sem tervezte meg, hanem evolúció útján fejlődött ki.” (Crick,
1988, 36.)
Amikor ezeket a nyilatkozatokat olvastam, felmerült bennem az a
kérdés, hogy vajon én egy természettudományt művelek-e, amikor
biológiával foglalkozom, vagy valamilyen ideológia területére
tévedtem? Milyen természettudomány az, amely arra kényszerít, hogy
amit látok, azt felejtsem el, és helyette higgyem el azt, amit X és Y
szaktekintély állít? El ne felejtsük már, hogy az 50-es években az
összes kelet-európai egyetemen − nekem is − azt tanították, hogy
kromoszómák nincsenek, mert azok a sejt fixálásának melléktermékei.
Megtévesztő dolog, hogy Francis Crick Nobel-díjasként nyilatkozik az
evolúcióról, amellyel kutatóként soha sem foglalkozott.
Az ilyen típusú állásfoglalások hamar elnyerik méltó büntetésüket.
Robert Laughlin fizikus, aki megszokta, hogy mindent meg lehet mérni,
és ki lehet számítani, ráadásul a kozmológia és az élet
keletkezésének fizikai feltételeit kutatja, azt írja nemrég
megjelent könyvében: „Jelenlegi biológiai tudásunk nagy része
ideológiai jellegű. Az ideológiai gondolkodás egyik fő tünete az
olyasféle magyarázat, amelyből nem vonhatók le következtetések, és
amely nem tesztelhető vagy ellenőrizhető. Az ilyen logikai
zsákutcákat antielméleteknek nevezem, mivel hatásuk éppen ellentétes
a valódi elméletekével: nem serkentik, hanem éppenséggel gátolják a
gondolkodást. Például a természetes szelekció általi evolúció,
amelyet Darwin fogalmazott meg mint nagy elméletet, manapság inkább
antielméletként működik, és arra használják, hogy elleplezzék a
zavarba ejtő kísérleti hibákat és hiányosságokat, és törvényesítsék a
megkérdőjelezhető felfedezéseket.” (Laughlin, 2005, 168–169.)
Félreértés ne essék: Robert Laughlin nem azt mondja, hogy Darwin
elmélete rossz, hanem azt, hogy rosszul használják, és ebben igazat
kell, hogy adjak neki.
Ugyancsak elgondolkoztató Sir Fred Hoyle, asztrofizikus és
matematikus véleménye: „Ugyan kiderült, hogy a darwini elmélet nem
lehet helyes teljes általánosságban, bennem mégis maradt egy kétely,
mert nehéz elhinni, hogy az elmélet egészében helytelen. Ha egy
elmélet megfigyeléseken alapul, mint a darwini elmélet, általában,
legalább a megfigyelések tartományán belül, érvényes marad. Baj akkor
van, ha az elméletet kiterjesztik a megfigyelések tartományán túlra.
Tehát felmerült a kérdés, hogy meddig érvényes egy elmélet, és miért
válik érvénytelenné egy ponton túl.” (Hoyle, 1987, 7.)
Az én meggyőződésem is az, hogy a neodarwinizmus és a materialista
világnézet pusztán praktikussági okokból olyan feltételezésekkel
terheli meg a darwini elméletet, amelyek magyarázatára az nem
alkalmas. Sir Fred Hoyle írja a továbbiakban: „A józan ész azt
sugallja, hogy a darwini elmélet helyes kis léptékben, de helytelen
nagy léptékben. A nyulak más, egy kicsit eltérő nyulaktól származnak,
de nem az őslevestől vagy a burgonyától. Az, hogy végső soron honnan
származnak, továbbra is megoldatlan probléma marad, ahogy sok minden
más a kozmoszban.” (Hoyle, 1987, 8.)
Az utóbbi években, és főleg a matematika, fizika és informatika
területéről jelentkező kritikai észrevételek hatására Richard Dawkins
is óvatosabbá vált, amit 2003-ban megjelent írásában tett nyilatkozata
tükröz: „Lehet, hogy Darwin diadalmaskodott a huszadik század végén,
(értsd: Dawkins diadalmaskodott Darwin nevében, a szerző megj.) de
tudomásul kell vennünk annak a lehetőségét, hogy új tények kerülnek
napvilágra, amelyek arra kényszerítik majd a huszonegyedik századi
utódainkat, hogy elvessék, vagy a felismerhetetlenségig módosítsák a
darwinizmust.” (Dawkins, 2003, 81.) Azt hiszem, hogy itt Dawkins
helyesen látja, hogy a 21. század tudománya ebben az irányban halad.
Nem érdektelen megismerni azoknak a molekuláris biológusoknak a
véleményét sem, akik az utolsó évtizedek legnagyobb horderejű
kutatásában, az emberi génanyag feltérképezésében vettek részt.
Francis Collins (Az Emberi Genom Program igazgatója): „Megkapó és
döbbenetes dolog ráébrednem, hogy először nyertünk bepillantást saját
használati útmutatásunkba, amelyet korábban csak Isten ismert.”
Gene Myers: (Celera Genomics, Marylandi Központ számítógépes
szakembere, aki részt vett az emberi genom feltérképezésében.):
„Elragadóan komplexek vagyunk molekuláris szinten… Ami valóban
bámulatba ejt, az élet architektúrája… a rendszer rendkívül komplex.
Olyan, mintha valaki megtervezte volna… Mérhetetlen intelligenciával
állunk itt szemben. Nem gondolom, hogy ez tudománytalan.”
Mialatt elolvastam egy kisebb könyvtárnyi irodalmat az evolúcióról,
lépten-nyomon szembekerültem azzal a kérdéssel, hogy mi tekinthető
tudománynak, és mi nem. Az a meggyőződés kezdett kialakulni bennem,
hogy miközben a tudósok nagy erőfeszítéseket tesznek annak érdekében,
hogy Istent kiszorítsák a tudományból meg a természetből, a tudomány
talán még soha sem állt olyan közel ahhoz, hogy Isten létét
bebizonyítsa, mint napjainkban. Nem lehetetlen, hogy a tudomány nem
fog tudni ellenállni annak a kísértésnek, hogy ezt a saját
eszközeivel meg is tegye. Örülök annak, hogy neves nagy tudósok −
legalább nyugdíjas korban − amikor már nem kell félniük attól, hogy
pályázataikat világnézeti alapon elutasítják, nem látnak gondolati
konfliktust a tudomány magas szintű művelése és az istenhit között.
Sir John Houghton a Royal Society tagja, az Oxfordi Egyetem fizikus
professzora, meteorológus írja: „Tudományunk Isten tudománya. Ő
viseli a felelősséget az egész tudományért. Az Univerzum tudományos
leírásában lévő figyelemre méltó rend, következetesség, megbízhatóság
és bámulatos komplexitás az Isten munkálkodásában lévő rend,
következetesség, megbízhatóság és komplexitás tükröződése.” (Houghton,
1995, 59.)
Sir Ghillean Prance botanikus, a világ két legnagyobb élő
növénygyűjteményének, a New York Botanical Gardennek és a Royal Kew
Gardensnek volt igazgatója, valamint az Amazonas őserdeinek világhírű
kutatója. Vele mindkét minőségében volt alkalmam találkozni és
beszélgetni. Nem tudtam, hogy hívő ember, mert mindannyiszor konkrét
tudományos kérdésekről cseréltünk eszmét, és ilyesmi nem került szóba
köztünk. Az Isten és a tudósok (Prance, 1997) című könyvben így
vall: „Régóta hiszem, hogy Isten az egész természet nagy Tervezője… A
tudományban szerzett összes tapasztalatom megerősítette hitemet. A
Bibliát úgy tekintem, mint a tekintély legfőbb forrását.”
Magam mint botanikus, az MTA és a Londoni Linné Társaság tagja vagyok.
Ötven éve tanítom a növényvilág fejlődéstörténetét, és tanúja voltam a
hatalmas fejlődésnek, ami fél évszázad alatt ezen a tudományterületen
végbement. Négy kontinensen kutattam a zárvatermő növényeket, és
térképeztem a növénytakarót. Több mint félszáz növénycsaládból több
mint félezer növényfajt fedeztem fel a tudomány számára. Mindezt nem
dicsekvésből írom, hanem hogy dokumentáljam hozzáértésemet a
kérdésekhez, amelyeket írásomban érintettem. Tapasztalataimról egy
interjúkötetben így vallottam: „A tudomány felelőssége, hogy
felmutassa a természet működésében a Teremtő törvényeit. Például azt,
hogy az evolúció az élővilág fejlődése; egy gazdálkodó rendszer
tökéletesedése, amelynek célja, hogy az élet legfontosabb funkciói: a
lét- és fajfenntartás egyre kevesebb anyagból, egyre jobb hatásfokkal
és egyre biztonságosabb módon teljesüljenek.” (Chikán, 2005, 34.)
Darwinnal szemben nincs bennem elutasítás, de meg kellett mutatnom,
hogy az én szakterületem ismeretanyagai hol és mennyit igazolnak a
természetes szelekció és a véletlenszerű fejlődés elméletéből. És
javára írom azt, hogy A fajok eredete második kiadását ezzel a
mondattal zárta: „Nagyszerűség van ebben a felfogásban, amely szerint
a Teremtő az életet a maga különböző erőivel eredetileg csak néhány
vagy csak egyetlen formába lehelte bele; és mialatt bolygónk a
nehézkedés megmásít-hatatlan törvénye szerint keringett, ebből az
egyszerű kezdetből végtelen sok szépséges és csodálatos forma
bontakozott ki és bontakozik ki ma is”.
Csalódott materialisták azonnal elkezdték terjeszteni, hogy ezt
Darwin a felesége kérésére írta. Akinek van egy csöpp emberismerete,
az öregember konok arcát látva aligha gondolhatja komolyan, hogy egy
nő kérésére hajlandó lett volna világraszóló felfedezését a
Teremtővel „megosztani”.
Kulcsszavak: növényvilág, evolúció, koevolúció, szakaszos fejlődés,
kreatív és szelektív, darwini és nem-darwini szakaszok, irány,
hajtóerő, stratégiák
IRODALOM
Borhidi Attila (2002):
Gaia zöld ruhája. Magyarország az ezredfordulón. MTA Stratégiai
Kutatások, Budapest
Borhidi Attila (2008): A
zárvatermők rendszertana molekuláris filogenetikai megközelítésben.
PTE, Pécs
Chikán Ágnes (2005):
Levelek a tudás fájáról. Agroinform, Budapest
Conway Morris, Simon
(1998): The Crucible of Creation. Oxford University Press, Oxford
Crick, Francis (1988):
Lessons from Biology. Natural History. 97, 36–38.
Darwin, Charles (1958): A
fajok eredete. Gondolat, Budapest
Darwin, Charles (1996):
Origin of Species. World Classics Edition. Oxford University Press,
Oxford
Dawkins, Richard (1986):
The Blind Watchmaker. Longmans, London
Dawkins, Richard (2003): A
Devil’s Chaplain. Weidenfeld and Nicholson, London
Eldredge, Nils (1985):
Time Frames: The Evolution of Punctuated Equilibria. Princeton Univ.
Press.
Gould, Stephen Jay (1977):
Evolution’s Erratic Pace. Natural History. 86, 5, 12-16.
Heribert-Nilson, Nils
(1952): Synthetische Artbildung. Fischer, Stuttgart–Jena
Houghton, John (1995): The
Search for God – Can Science Help? Lion, Oxford
Hoyle, Fred (1987):
Mathematics of Evolution. Weston Publ. Cardiff University Press
Laughlin, Robert (2005): A
Different Universe: Reinventing Physics from the Bottom Down. New York
Basic Books
Maynard-Smith, John –
Szathmáry Eörs (1995): The Major Transitions in Evolution. Freeman,
Oxford–New York
Maynard-Smith, John –
Szathmáry Eörs (2002): A földi élet regénye. Vince, Budapest
Podani János (2003): A
szárazföldi növények evolúciója és rendszertana. Eötvös Kiadó, ELTE
Prance, Sir Ghillean
(1997): In: Mike Poole (ed.): God and the Scientists. CPO, Worthing
Raup, David (1979):
Conflicts between Darwin and Paleontology. Bulletin. Field Museum of
Natural History, Chicago
Valentine, James (2004):
On the Origin of Phyla. Univ. Press, Chicago
|
|