|
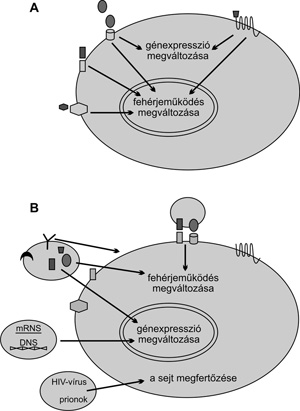
A fehérjéken és lipideken kívül néhány
mikrovezikulában nukleinsavakat, mRNS-t, miRNS-t és DNS-t is
azonosítottak. Mi több, a fertőző partikulumok terjedésének egy
újonnan felismert formája is összefügghet a mikrovezikulákkal,
amennyiben HIV-vírust és prionokat is találtak már bennük.(Valadi et
al., 2007; Ratajczak et al., 2006)
Minden valószínűség szerint a MV-k membránjában és
citoplazmájában jelen lévő szénhidrátoknak is fontos szerepük van az
intercelluláris kommunikációban, ám ezek vizsgálata napjainkban még
gyerekcipőben jár.
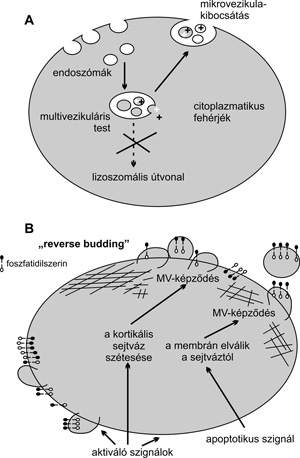
A mikrovezikulák mérése, vizsgálhatósága
Izolálás • A MV-k izolálását sejtmentes biológiai mintákból,
leggyakrabban ultracentrifugálás módszerével végzik. A különféle
munkacsoportok izolálási módszerei, az alkalmazott izoláló közeg,
ülepítési idő és sebesség, eltérő.
Morfológiai vizsgálatok • A tisztított (izolált)
MV-kat kicsiny méretük miatt adekvát módon elektronmikroszkópos
módszerrel lehet láthatóvá tenni. Az alkalmazott elektronmikroszkópos
mintaelőkészítés minden esetben az aktuális vizsgálati rendszertől és
kérdésfeltevéstől függ.
Áramlási citometria (flow cytometria, FACS) • Az
áramlási citometria a MV-k vizsgálatának legelterjedtebb és egyik
legelfogadottabb módszere. Ismert számú standard méretű plasztikgyöngy
felhasználásával a módszer lehetővé teszi a biológiai minták
MV-számának és méretbeli megoszlásának meghatározását. A MV-k
sejtfelszíni fehérjemintázatának immunfenotipizálással történő
azonosítása lehetővé teszi eredetük meghatározását, de hozzásegít a
funkcionális jellemzésükhöz és célsejtjeik azonosításához is.
Korlátozza a MV-k FACS-analízisét az, hogy a konvencionális készülékek
a 300 nm-nél kisebb partikulumok elkülönítésére nem képesek.
Funkcionális vizsgálatok • Az izolált MV-k által
közvetített biológiai hatások vizsgálatára leggyakrabban in vitro
sejtkultúrákat használnak fel. Ezek összeállítása, a hatás időtartama
minden esetben a kérdésfeltevéstől függ.
A mikrovezikulák szerepe
A mikrovezikulák kifejthetik hatásukat az őket kibocsátó anyasejtek
közvetlen környezetében, de hatással lehetnek viszonylag távoli
sejtekre is, hiszen a vérben keringve a szervezetben bárhová
eljuthatnak. Irodalmi adatok szerint a vérplazmában 5–50 µg/ml
koncentrációban fordulnak elő, eredetüket tekintve főként
fehérvérsejt, vörösvértest, vérlemezke és endotél eredetűek, de
speciális fiziológiás és patofiziológiás állapotokban más sejtekből
származók is kimutathatók (például a terhesség során megjelennek a
méhlepény sejtjeiből, a trofoblasztokból származó mikrovezikulák is).
Patológiás állapotokban mennyiségük és összetételük megváltozik,
például gyulladásban, tumorképződéskor, szív- és érrendszeri
betegségekben, veszélyeztetett terhességben (preeclampsia, koraszülés)
számuk jelentősen megemelkedik.
Általánosságban igaz, hogy a mikrovezikulák
pleiotróp hatásúak, vagyis ugyanazon sejtből származó mikrovezikulák
több különféle sejttel is kapcsolatba léphetnek, jóllehet ez a kémiai
kötődés természetesen nem véletlenszerű, hanem a fogadó sejt és a
mikrovezikula membránfehérje-mintázatától függ. A MV–célsejt kapcsolat
nyomán a fogadó sejtben egyszerre akár többféle szignalizációs kaszkád
is elindulhat, amely hatásában egyaránt lehet serkentő és gátló. Az
antigénprezentáló sejtekből származó MV-k például T-sejt aktivációt
indukálnak, míg a tumorsejt vagy trofoblaszt eredetű mikrovezikulák
immunszuppressziót váltanak ki. A funkcióban bekövetkező változások
mellett elképzelhető azonban az is, hogy a MV új tulajdonságokkal
ruházza fel a sejtet – új fenotípust kölcsönöz neki – azáltal, hogy
membránja beépül a célsejt membránjába.
A MV-k hatásmechanizmusának minél pontosabb
feltérképezése új diagnosztikus és prognosztikus markerek
felfedezését, illetve új terápiás célpontok kifejlesztését teszi
lehetővé (Andreola et al., 2002; Redman – Sargent, 2008; Freyssinet,
2003; Pap et al., 2009).
Mikrovezikulák szerepe terhességben
Kutatócsoportunk néhány év óta foglalkozik a MV-k anya–magzat
kommunikációban betöltött szerepével, ezért e fejezetben a MV-k és a
terhesség kapcsolatát kicsit bővebben, saját eredményeinkkel kibővítve
tárgyaljuk.
Habár immunológiai értelemben az anyai szervezet
számára a magzat idegen (szemiallograft), hiszen genetikai
állományának fele az apától, fele az anyától származik, jelen tudásunk
szerint a terhesség zavartalan lefolyásának alapvető feltétele az
idegen magzati antigének anyai felismerése és az ennek következtében
kialakuló anyai immunválasz (Szekeres-Barthó, 2005).
A mikrovezikulák által közvetített sejtszintű
kommunikációs útvonal terhességben játszott szerepéről még kevés
információ áll rendelkezésünkre, jelenlétüket azonban kimutatták
terhes nők vérében, sőt a magzatvízben is. Habár biológiai
jelentőségük mind a mai napig nem teljesen tisztázott, az mindenesetre
körvonalazódni látszik, hogy kiemelkedő szerepük van az immunrendszer
működésének szabályozásában. Az anyai vérben fenotípus-vizsgálatokkal
többek között vérlemezkékből származó, sőt méhlepény eredetű MV-k
jelenlétét is kimutatták. A klinikai állapottal történő összevetés
szerint szövődménnyel járó terhességben (preeclampsia) a
vérlemezkékből származó MV-k mennyisége emelkedik (Vanwijk et al.,
2002).
Mivel a MV-k sejtfelszíni fehérjestruktúrájuk
és/vagy a citoplazmájukban szállított anyagaik révén képesek
befolyásolni a sejtműködést, ezért feltételezhető, hogy az anya és a
magzat közti aktív kommunikációban is szerepet játszanak. Hipotézisünk
szerint egyaránt befolyással lehetnek az anyai immunrendszer és a
méhlepény működésére is. Kísérleti munkánkban szövődménymentes
terhesek perifériás vérében detektáltuk a keringő MV-mintázatot, és
megvizsgáltuk a keringő MV-k immunmoduláló hatását.
Eredményeink szerint terhességben a keringő MV-knak
mind a mennyisége, mind az összetétele megváltozik. A vérlemezkékből
származó MV-k mennyisége lecsökken, ugyanakkor a nem specifikus
immunválasz közvetítésében fontos szerepet játszó monocitákról leváló
részecskék száma megnő, és nem utolsósorban jelen vannak magzati
sejtekből származó MV-k is. Sikerült tehát igazolnunk, hogy
terhességben megváltozik a perifériás vérben keringő MV-mintázat. A
MV-k által közvetített hatások vizsgálata során kimutattuk, hogy a
magzati sejtekből és az anyai vérlemezkékből származó MV-k egyaránt
képesek kötődni az anyai T-limfocitákhoz, és ez a kapcsolódás
megváltoztatja ezeknek a sejteknek a működését (Pap et al., 2008; Kiss
et al., 2008).
Összességében tehát igazoltuk, hogy a terhességre
jellemző anyai immunválasz fenntartásában és szabályozásában jelentős
szerepük van a MV-k által közvetített kommunikációs útvonalaknak is.
Kulcsszavak: mikrovezikula, sejtek közötti kommunikáció, terhesség
IRODALOM
Andreola, Giovanna – Rivoltini, L. –
Castelli, C. – Huber, V. – Perego, P. – Deho, P. – Squarcina P. –
Accornero P. – Lozupone F. – Lugini L. – Stringaro A. – Molinari, A. –
Arancia, G. – Gentile, M. – Parmiani, G. – Fais, S. (2002): Induction
of Lymphocyte Apoptosis by Tumor Cell Secretion of Fasl-Bearing
Microvesicles. The Journal of Experimental Medicine. 195, 10,
1303–1316.
Coleman, Mathew L. – Sahai, EA. – Yeo, M.
– Bosch, M. – Dewar, A. – Olson, MF. (2001): Membrane Blebbing During
Apoptosis Results from Caspase-Mediated Activation of ROCK I. Nature
Cell Biology. 3, 4, 339–345.
Distler, Jüngel H. – Pisetsky, D. S. –
Huber, L. C. – Kalden, J. R. – Gay, S. – Distler, O. (2005):
Microparticles As Regulators of Inflammation: Novel Players of
Cellular Crosstalk in the Rheumatic Diseases. Arthritis & Rheumatism.
52, 11, 3337–3348.
Freyssinet, Jean-Marie (2003): Cellular
Microparticles: What Are They Bad Or Good for? Journal of Thrombosis
and Haemostasis. 1, 7, 1655–1662.
Johnstone, Rose M. (2006): Exosomes
Biological Significance: A Concise Review. Blood Cells, Molecules and
Diseases. 36, 2, 315–321.
Kiss Attila András – Pap E. – Falus A. –
Pállinger É. (2008): Mikrovezikulumok immunológiai szerepe az
anya–magzat kommunikációban. Magyar Nőorvosok Lapja. 71, 269–276.
Pap Erna – Pállinger É. – Falus A. – Kiss
A. A. – Kittel A. – Kovács P. – Buzás E. I. (2008): T Lymphocytes Are
Targets for Platelet- and Trophoblast-Derived Microvesicles During
Pregnancy. Placenta. 29, 9, 826–832.
Pap Erna – Pállinger É. – Pásztói M. –
Falus A. (2009): Highlights of a New Type of Intercellular
Communication: Microvesicle-Based Information Transfer. Inflammation
Research. 58, 1–8.
Ratajczak, Janina – Wysoczynski, M. –
Hayek, F. – Janowska-Wieczorek, A. – Ratajczak, MZ. (2006):
Membrane-Derived Microvesicles: Important and Underappreciated
Mediators of Cell-to-Cell Communication. Leukemia. September 2006. 20,
9, 1487–1495. Epub 20 July. Review.
Redman, Christopher W. – Sargent, Ian L.
(2007): Microparticles and Immunomodulation in Pregnancy and
Pre-Eclampsia. Journal of Reproductive Immunology. 76, 1–2, 61–7.
Redman, Christopher W. – Sargent, Ian L.
(2008): Circulating Microparticles in Normal Pregnancy and
Pre-Eclampsia. Placenta. 29, S1, 73–77.
Szekeres-Barthó Júlia (2005): A terhesség
immunogenomikai vonatkozásai. Magyar Tudomány. 6, 708–713.
Théry, Clotilde – Zitvogel, L. –
Amigorena, S. (2002): Exosomes: Composition, Biogenesis and Function.
Nature Rewievs Immunology. 2: 569–579.
Valadi, Hadi – Ekström, K. – Bossios, A. –
Sjöstrand, M. – Lee, JJ. – Lötvall, JO. (2007): Exosome-Mediated
Transfer of Mrnas and Micrornas Is a Novel Mechanism of Genetic
Exchange between Cells. Nature Cell Biology. 9, 6, 654–659.
Vanwijk, Marja J. – Nieuwland, R. – Boer,
K. – Van Der Post, J. A. M. – Vanbavel, E. – Sturk, A. (2002):
Microparticle Subpopulations Are Increased in Preeclampsia: Possible
Involvement in Vascular Dysfunction? American Journal of Obstetrics &
Gynecology. 187, 450–456.
|