|
|
Az utóbbi néhány
évtized pszichológiájában, részben párhuzamosan a molekuláris
biológia ötven éve zajló forradalmi átalakulásával, részben azonban
saját belső okokból is, igen megerősödött a genetikai mozzanatok
iránti érdeklődés. Ennek a gondolatmenetnek vannak olyan változatai,
amelyek a genetikában keresnek fogódzót az emberi elme egyetemes
egységes szerveződésének megragadására, más felfogások azonban azt
hangsúlyozzák, hogy a genetika leglényegesebb üzenete a lelki vonások
tekintetében is meglévő variabilitás kiemelése. Az
1. táblázat
mutatja a pszichológiai elmélet genetikai érdekességű
megújulásának legfontosabb képviselőit.
A mai evolúciós pszichológia által oly sokat
kritizált klasszikus társadalomtudomány, akárcsak a klasszikus
tanuláselmélet, kumulatív tanulógépezetnek képzelte el az embert,
akinek vonásai mintegy összegződve hozzák létre pillanatnyi tudását és
pillanatnyi érdeklődéseit és így tovább. Az utóbbi néhány évtized
fejlődése az embert bonyolult információfeldolgozó lényként, majd
bonyolult társas konstruáló lényként értelmezi. A kognitív
pszichológia kibontakozása természetesen oda vezetett, hogy az
örökléselvű meghatározottságot hangsúlyozó szerzők már nem egy üres
szervezetet elképzelő behaviorizmussal vitáznak, hanem egy igen
kifinomult elképzeléssel, mely különböző biológiai
konstrukcionizmusok formájában működik. Mindkét felfogás az emberi
megismerés és érzésvilág általános építményét, elegánsan azt szoktuk
mondani, hogy a kognitív és az affektív architektúrát illetően
fogalmaz meg elképzeléseket. A preformista felfogás, amelynek
legjelentősebb képviselői Noam Chomsky és például Jerry Fodor, azt
hangsúlyozza, hogy minden egyetemesen megjelenő megismerési és
szociális mozzanat genetikailag meghatározott. Evolúciós örökségünk,
mondják sokan, határozza meg, hogy éppen ilyenné váltunk, vagyis azt,
hogy mi emberek különlegesen hajlunk szerkezetszerű tanulási elveket
követni és így tovább. Az összefoglaló néven környezetelvű biológiai
konstrukcionizmusnak nevezhető felfogások viszont azt hangsúlyozzák,
hogy az egyéni elsajátítás mechanizmusa során rögzülnek nemcsak az
egyedi tudások vagy az egyedi preferenciák, hanem a megismerési
architektúra szerkezetei is. Biológiai adományaink csak sajátos
különleges tanulási elvek és nem kész tudások. Ennek a felfogásnak
különböző változatai vannak, jellegzetes képviselői Anett
Karmiloff-Smith, Michael Tomasello, Magyarországon pedig Gergely
György és Csibra Gergely.
A különböző típusú
genetikai felfogásokat hangsúlyozó elképzelések világában kitüntetett
jelentőséget kap nemcsak az általános vonások evolúciós örökségként és
genetikai meghatározottságként értelmezése, hanem az egyéni
különbségek problémái is. A molekuláris genetika fejlődésével ennek
vizsgálata a klasszikus módszerek (ikerkutatás, örökbefogadás) mellett
egyre többször kísérel meg tényleges leképezést a genomváltozatok és a
kiugró viselkedésváltozatok között. A
2. táblázat néhány jellegzetes
példát mutat arra, milyen kérdéseket szoktak ezen a téren kutatni.
(Pszichiátriai vonatkozásaira jó összefoglaló Burmeister et al.,
2008.) A genetikai eredetű fejlődési variációk és a valódi mentális
betegségek kutatása sajátos disszociációs logikából indul ki. Ez a
disszociációs logika a klasszikus genetikára épít: vannak gének,
amelyek meghatározott azonosítható tulajdonságokat szabályoznak. A
felfogás másik támpontja szerint az emberi megismerés építménye is
moduláris felépítésű, hasonlít a genom felépítéséhez, egy-egy
területre, megismerésmódra (például: arcokra, szavakra stb.)
specializálódott alrendszerekből áll, és ezeknek az alrendszereknek a
sérülése felel meg valamilyen gén sérülésének. Ennek a disszociatív
logikának a másik szempontja a klasszikus genetika mellett a
klasszikus neuropszichológia. Feltételezi, hogy ugyanolyan módon,
ahogy egy agyvérzés vagy baleset következtében kiiktatódik például a
Wernicke-terület, és ennek megfelelően zavarok lépnek fel a beszéd
megértésben, illetve a szókincs szerveződésében, ugyanez lesz érvényes
a fejlődésre is. A kiindulás tehát az, hogy egy jellegzetes
gén-agy-megismerési modul-megfelelés képzelhető el.
Egy példát fogok bemutatni arra, hogy ezt még az
igen jól körvonalazott genetikai zavarok esetén is mára sokkal
árnyaltabban értelmezzük. Úgy véljük, hogy genetikai értelemben minden
magasabb szintű viselkedés poligénes meghatározottságú, és ennek
megfelelően zavarai is számos tényező zavarát tükrözik, másrészt
feltételezzük, hogy a megzavart fejlődés is jellegzetes idői
mintázatban, fejlődési mechanikával bontakozik ki. Amikor a genetikai
zavar például fejlődési elmaradást eredményez, ennek következtében
hosszú évek alatt előállhat egy jellegzetes architekturális kép.
Példa: a Williams-szindróma
A Williams-szindróma egy közel fél évszázaddal ezelőtt azonosított
jellegzetes fejlődési zavar, amely a 7. kromoszóma egy
mikrodeléciójával kapcsolatos. Ez a mikrodeléció a 7. kromoszóma
hosszú karján, ma már tudjuk, hogy meglehetősen sok, összesen kb. húsz
gént érint, és magában a genetikai zavarban is sokkal nagyobb egyéni
változatok vannak, mint azt a kutatás indulásakor gondoltuk (Morris –
Mervis, 2000).
Kiinduláskor a következő jellegzetes mintázatot
figyelték meg a Williams-szindrómával jellemzett személyeknél (Bellugi
et al., 2000; Lukács, 2005):
• az elasztin enzim hiánya miatt igen erős
keringési és vesezavarok figyelhetők meg;
• jellegzetes arcformájuk van;
• az IQ az enyhe fogyatékos régióban van, 50 és 80
között;
• igen erősek a téri orientáció zavarai;
• jellegzetes számolási zavarok figyelhetők meg;
• ezzel párhuzamosan viszonylag jók a nyelvi
készségek;
• igen szociábilisak.
A kognitív zavarok fejlődése iránt érdeklődő
„pszichogenetikusok” számára a Williams-szindróma azért volt különösen
érdekes és fontos, mert jellegzetes disszociációs párokat képzeltek el
például Steven Pinker (2004) vagy Ursula Bellugi és munkatársai
(2000):
• rossz téri tájékozódás – jó nyelvi készségek
• a nyelven belül viszonylag jó nyelvtan – furcsán
szerveződő szókincs
• jó szociábilitás – alacsony szintű végrehajtó
működések.
A Magyar Williams-szindróma Kutatás keretében
mintegy tíz éve vizsgáljuk ezeket a
felmerült disszociációkat. Ennek a nyelv szempontjából sajátos
érdekességét az adja, hogy a Williams-szindrómásokra vonatkozó adatok
javarészt indoeurópai nyelvekből származnak, például az a mintázat,
amelyet Steven Pinker (1991) és Harald Clahsen (1999) hirdettek, azt
emeli ki, hogy a Williams-szindrómásoknak furcsa szerveződésű lenne a
szókincsük, ugyanakkor viszonylag jó,
érintetlen lenne a nyelvtani készségük. Ezzel szemben mi sokkal
variábilisabb mintázatokat találunk (Kovács, 2004; Lukács, 2005;
Lukács et al., 2005) Vannak olyan esetek, amint a
3. táblázat
mutatja, ahol valódi minőségi disszociáció van; vannak olyan esetek,
amikor viszont lassulás van nagy egyéni eltérésekkel; és vannak olyan
esetek, ahol a tipikus fejlődéssel azonos mintázatot kapunk.
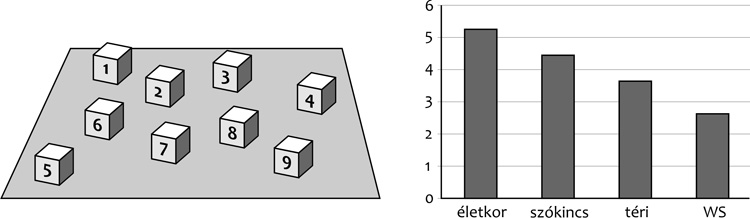
Egy jellegzetes példát mutatok be a disszociációra, a téri
megismerést, mint az
1. ábra
mutatja. A Williams-szindrómás csoportban tizenkét év körüli gyerekek
vettek részt. Az ún. téri kontroll viszont 3 és fél éves életkorú
gyerekcsoport volt. A Williams-szindrómások még náluk is jóval
gyengébb teljesítményt mutatnak vizuális mintázatok felidézésében a
Corsi-próbában.
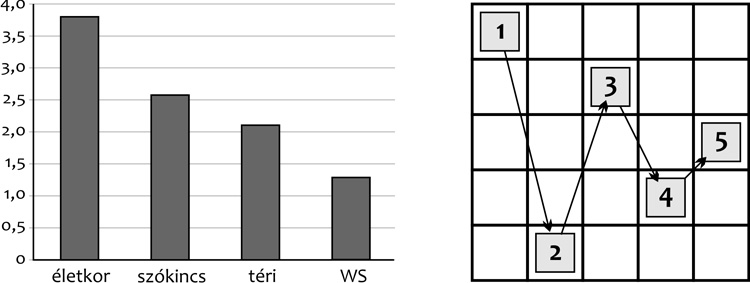
Racsmány Mihály (2004, Racsmány et al., 2002) azt
is kimutatta, hogy egy bonyolultabb helyzetben, amikor meg kell
tanulni egy vizuális kopogási mintázatot,
szintén megfigyelhető a Williams-szindrómások igen korlátozott
téri teljesítménye. (2.
ábra)
Kutatásunkban igen sok vizsgálatot végeztünk a
Williams-szindrómások nyelvi profiljára nézve. Számos helyen
beszámoltunk erről (Lukács et al., 2005) ezért csak néhány példát
mutatunk be arra, hogy a klasszikus elképzelésekkel szemben a
Williams-szindrómások sok tekintetben azonosan működnek a tipikus
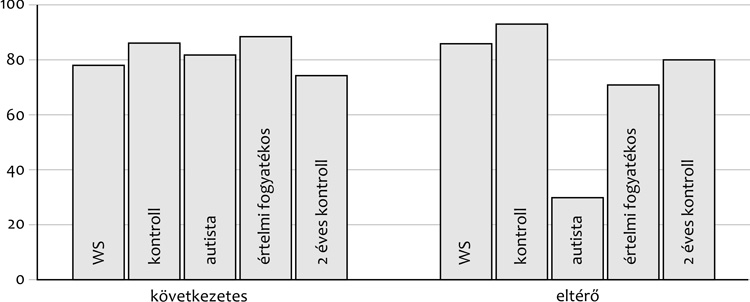
fejlődésűekkel, illetve lassabbak. A
3. ábra azt mutatja be, hogy a
szótanulás közbeni társas támpontok felhasználása során a
Williams-szindrómások tipikusan ugyanúgy viselkednek, mint a különböző
kontrollgyermekek, szemben az autistákkal, akik számára nehéz
megtanulni azt, hogy a kísérletvezető és
a saját tekintetük eltérően irányul tárgyakra szótanulási helyzetben.
Sajátosan érdekes, mert a nemzetközi
szakirodalommal sok vita tárgya, hogy vajon a Williams-szindrómásoknál
tényleg igaz-e az, hogy a ragozási kivételeket furcsán kezelnék, éppen
azért, mert szabályrendszerük jól fejlett lenne, míg a
szóstabilizáció-rendszerük, mely a kivételeket (hó-havat, ló-lovat
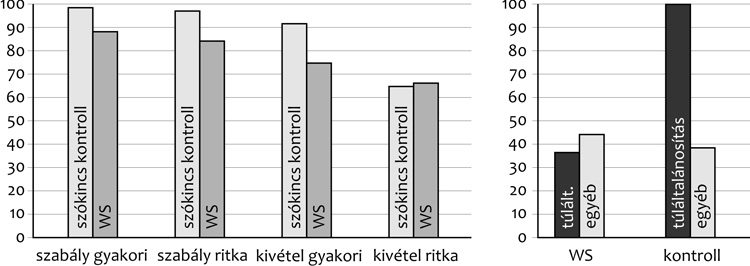
stb.) kezelné, sokkal fejletlenebb. Mint a
4. ábra mutatja, azt találtuk, hogy
a Williams-szindrómásoknál is ugyanúgy alakul a teljesítmény
mintázata, mint a szókincs alapján velük illesztett
kontrollgyerekeknél. Ráadásul, mint az ábra jobb oldala mutatja, ha
túláltalánosítási hibákat tekintünk, (sót, kenyért stb.), akkor a
túláltalánosítási hibák inkább az átlagos fejlődésű gyerekekre
jellemzőek. Vagyis a magyar nyelvi adatokat használva sikerült
rámutatnunk arra, hogy egy szokványosan, az indoeurópai adatok alapján
minőséginek, disszociatívnak tűnő fejlődés tekintetében nem érvényes a
disszociáció.
Különlegesen érdekes, hogy mi történik a téri
nyelvben. A Williams-szindrómával kapcsolatban a legkiugróbb kognitív
probléma a téri tájékozódás zavara. Azt találjuk, hogy a
Williams-szindrómás gyerekek (emlékeztetek a 2. ábrára)
gyakorlatilag képtelenek megjegyezni például két fordulót egy útvonal
kialakítása során. Vajon hogyan sajátítják el az olyan ragokat, mint
házban, házra, házról, vagy névutókat, mint ház
mögött, ház mögé, ház mögül stb.? Az 5. ábra
azt mutatja, hogy mint az a téri teljesítményük alapján
várható is, a Williams-szindrómások sokkal gyengébb teljesítményt
mutatnak, mint a szókincs kontrollok, akik szinte százszázalékos
teljesítményt mutatnak. (A kísérletben tizenkét éves
Williams-szindrómás gyerekeket hasonlítunk össze hatéves átlagos
fejlődésűekkel.) A forrás-kifejezések számukra a legnehezebbek.
Mindannyiunk számára nehezebb a házból, mint a házba, és
a legkönnyebb a házban.
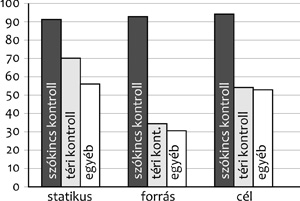
5. ábra
• A Williams-szindrómások téri nyelv-használata hasonló
teljesítményt mutat, mint a tipikus fejlődésű gyerekeké (Lukács, 2004) |
|
|
Lukács Ágnes (2004) kutatásaiban azt is kimutatta,
hogy ha olyan használatait vetjük össze a téri ragoknak, ahol azok hol
tériek (az oroszlán megszökött a ketrecből), hol pedig elvontak és
mentálisak (Pisti tanult a balesetből) akkor meglepő módon a
Williams-szindrómásokra is igaz az, hogy a téri jelentést könnyebben
kezelik. Itt azonban kicsit óvatosnak kell lennünk, mert a téri
jelentés könnyebbsége egy olyan felidézési helyzetben jelenik meg,
ahol a gyerekeknek nem kellett értelmezniük a téri elrendezést. De
mindenesetre ez is arra utal, hogy még a legérzékenyebb nyelvi
területen, a téri nyelv elsajátításában is ugyanazt a mintázatot
követik, mint az átlagos gyerekek.
Számos olyan vizsgálat is van, amely az eredeti
disszociációs logika harmadik komponensét, az agyat is eléri. Azt
találják, hogy a Williams-szindrómások jellegzetes agyfejlődési zavara
a parieto occipitalis sulcus, a fali lebeny és a tarkólebeny
érintkezési területének alulfejlettsége. Ez a terület a téri
tájékoztatásért felelős parietális, és a vizuális teljesítmények
alapjaiért felelős tarkólebenyi területek alulfejlettségét
eredményezi. Tehát magában a téri kogníció alulfejlettségében
megtaláljuk a határozott megfelelést a kognitív disszociatív zavar, az
idegrendszeri zavar és a genetikai elváltozás között (Reiss et al.,
2004; Meier et al., 2006) Ugyanakkor a többi területen nincs meg ez a
jellegzetes viselkedéses disszociáció.
Mi a példa tanulsága?
A genetikai zavarok plasztikus kibontakozása
A genetikai zavarok értelmezését célzó mai kognitív kutatás az
eredeti, a szerzett agysérülések disszociatív logikáját és a
klasszikus genetikát felhasználó mindent vagy semmit megfontolások
helyett új érvelést állít előtérbe. Ennek lényege egyszerűen
megfogalmazható. Mára észrevettük, hogy a magasabb kognitív funkciókra
nem lehet egy klasszikus mendeli genetikai képet rávetíteni, amely
minden kognitív vonást egy sajátos gén általi szabályozottság alá
rendelne. Maga a genetika is sokkal plasztikusabbá vált.
Rácsodálkozunk arra, hogy a mintegy húszezer gén talán nem is tud
egyenként olyan sok mozzanatot meghatározni, ezért a kognitív fejlődés
genetikai meghatározottságában is előtérbe kell helyeznünk a
szabályozó működések jelentőségét.
Másrészt észrevesszük azt is, hogy a magasabb
kognitív rendszerekben számos olyan többkomponensű rendszer van,
amelyek közös adaptív funkciókat valósítanak meg. Nem mindent lehet a
színvakság analógiájára értékelni, ahol például vagy megvan, vagy
nincs meg a vörös-zöld rendszer. Sokszor a megfelelő analógia a
szürkületi és a nappali látás, ahol kétféle rendszer egyazon funkciót
a nagy megvilágítási körülmények között is érvényesülő vizuális
viselkedésirányítás valósítja meg (Kovács, 2004).
Harmadrészt, az idegrendszer bizonyos részei az
emlősöknél, abban a világban, ahol az agykéreg egy része hatalmas
tanulógépezetként működik, éppenséggel genetikailag meghatározottan
viszonylag hajlékony. Ebből az következik, hogy maguk a genetikailag
jelenségszinten minőségi eltérést mutató kognitív zavarok is fejlődési
úton érik el ismert képüket. Conrad Waddington (1957) híres
kanalizációs példáját, amely az evolúciósan kialakult tájkép és az
egyéni fejlődés útját próbálja meg bemutatni (6. ábra), ma
többdimenziós epigenetikus térképek váltják fel.
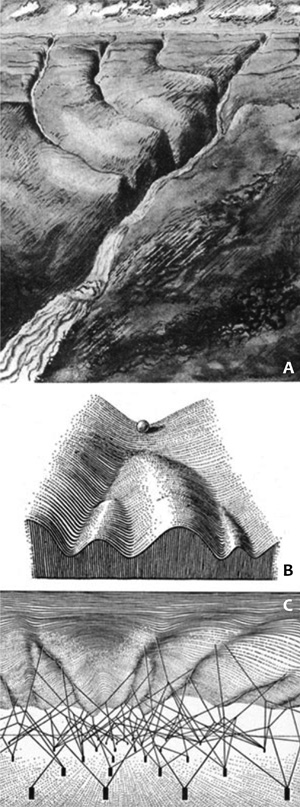
6. ábra
• Conrad Waddington (1957) elképze-lése a fejlődés útját
preferáltan alakító genetikai tájképről • a – valós tájkép; b – a
fejlődési tájkép • c – a mögöttes genetikai rendszer. A sötét hasábok
a gének, az összekötő vonalak a fenotípushoz vezető sokrétű oksági
láncok.
Ma úgy képzeljük el, hogy ebben a tájképben
lehetnek jellegzetes szakadékok, ezek felelnek meg az olyan drámai
genetikai alterációknak, mint a Williams-szindróma, lehetnek
meredekebb pályák, ezek felelnek meg a többgénes bonyolult
meghatározottságoknak, és lehetnek nagyobb lejtések, ezek felelnek meg
a fejlődési variációknak.
Mindebből egy olyan elképzelés bontakozik ki, amely szerint a szerzett
zavarok és a genetikai zavarok nem azonos logika szerint
értelmezendőek. Ma már nem tartható a korai elképzelés, amit például
Pinker hangsúlyozott (Pinker, 1991, 2004) az agysérülések és a
genetikai meghatározottság azonos logikájáról. Pinker felfogásában
egy olyan kép élt, ahol a Williams-szindrómának a Broca-terület épsége
és a Wernicke-terület kompromittáltsága, míg például a specifikus
nyelvi zavarnak a Broca-terület zavara és a Wernicke-terület
viszonylagos épsége felelne meg. Számos felfogás fogalmazódott meg az
utóbbi húsz évben, amely dinamikusabb képet hangsúlyoz Ilyen például
Michael S. C. Thomas és Anett Karmiloff-Smith (2002) elképzelése,
amely szerint a gondolkodás rendszerének kialakulása során bekövetkező
zavarok, vagyis a korai genetikai zavarok az egész tanulási
mechanizmust befolyásolják. De ilyen Elisabeth Bates (2004) felfogása
is, amely szerint a tanulási sebesség lelassulása mellett nincs meg
annyi kumulatív tapasztalat, amelyből ugyanúgy tudnának kibontakozni a
fejlődési rendszerek.
A mi felfogásunk szerint azok a mozzanatok a
Williams-szindrómában, amelyek nem minőségi különbségekre, hanem
lassabb tanulásra utalnak, azt eredményezhetik, hogy a lassabb tanulás
révén addig nem ér el a gyerek olyan fejlődési szintet, mint a tipikus
fejlődésű gyermek, amíg a tanulás kritikus ablakai le nem zárulnak.
Úgy gondoljuk, hogy a jövő útja itt a hagyományos
orvostudományban ismert genetikai szemlélet és az embriológiai
szemlélet összekapcsolása a viselkedés meghatározottságával. A zavart
fejlődés, legyen az betegség vagy súlyos változat, megértése
elősegítheti, hogy pontosabban tisztázzuk azt az elméleti kérdést,
hogy milyen finom szabályozású kritikus periódusai vannak a
fejlődésnek, s hogy ebben a szabályozásban milyen kulcspontokat
teremtenek vagy képviselnek a gének, s egyáltalán hol van beavatkozási
lehetőség egy olyan genetikai meghatározottságban, ahol a genetika egy
bonyolult tanulási rendszer beállítási mechanizmusaiban érintett.
Kulcsszavak: pszichogenetika, Williams-szindróma, téri
nyelv zavarai
IRODALOM
Bates, Elisabeth (23004): Explaining and
Interpreting Deficits in Language Development across Clinical Groups:
Where Do We Go from Here? Brain and Language. 88, 2, 248–253.
Bellugi, Ursula – Lichtenberger, L. –
Jones, W. – Lai, Z. (2000): The Neurocognitive Profile of Williams
Syndrome: A Complex Pattern of Strengths and Weaknesses. Journal of
Cognitive Neuroscience. 12, 7–29.
Burmeister, Margit – McInnis, M. G. –
Zöllner, S. (2008): Psychiatric Genetics: Progress Amid Controversy.
Nature Reviews Genetics. 9, 527–540.
Clahsen, Harald (1999): Lexical Entries
and Rules of Language: A Multidisciplinary Study of German Inflection.
Behavioural and Brain Sciences. 22, 991–1060.
Fisher, Simon E.(2006): Tangled Webs:
Tracing the Connections between Genes and Cognition. Cognition. 101,
270–297.
Kovács Ilona (2004): Visual integration:
Development and impairments. Akadémiai, Budapest
Lukács Ágnes (2005): Language Abilities in
Williams Syndrome. Akadémiai, Budapest
Lukács Ágnes – Pléh Cs. – Racsmány M.
(2005): Nyelvi képességek Williams-szindrómában. Pszichológia. 25,
309–347.
Lukács Ágnes – Pléh Cs. – Racsmány M.
(2007): Spatial Language in Williams Syndrome: Evidence for a Special
Interaction? Journal of Child Language. 34, 311–343.
Meyer-Lindenberg, Andreas – Mervis, C. B.
– Berman, K. F. (2006): Neural Mechanisms In Williams Syndrome: A
Unique Window to Genetic Influences on Cognition and Behaviour. Nature
Reviews Neuroscience. 7, 380–393.
Morris, Colleen A. – Mervis, Carolyn B.
(2000): Williams Syndrome and Related Disorders. Annual Review of
Genomics and Human Genetics. 1, 461–484.
Pinker, Steven (1991): Rules of Language.
Science. 253, 530–535
Pinker, Steven (2004): A nyelvi ösztön.
Typotex, Bp.
Pléh Csaba (2008): A pszichológia örök
témái. Typotex, Budapest
Pléh Csaba – Lukács Ágnes (2005):
Alkalmazkodás és plaszticitás a nyelv evolúciójában és egyedfejlődési
patológiájában. Magyar Tudomány. 111, 62–69.
WEBCÍM >
Racsmány Mihály (2004): A munkamemória
szerepe a megismerésben. Akadémiai, Budapest
Racsmány Mihály – Lukács Ágnes – Pléh
Csaba (2002): Munkamemória és nyelvelsajátítás Williams- szindrómában.
Pszichológia. 22, 255–266.
Reiss, Allan L. – Eckert, M. A. – Rose, F.
E. – Karchemskiy, A. – Kesler, S. – Chang, M. – Reynolds, M. F. –
Kwon, H. – Galaburda, A. (2004): An Experiment of Nature: Brain
Anatomy Parallels Cognition and Behavior in Williams Syndrome. Journal
of Neuroscience. 24, 5009–5015.
Thomas, Michael S. C. – Karmiloff-Smith,
Anett (2002): Modelling Typical and Atypical Cognitive Development.
In: Goswami, U. (ed.)> Handbook of Childhood Development. Blackwell
Publishers, London
Waddington, Conrad Hal (1957): The
Strategy of the Genes. George Allen & Unwin, London |
|