|
|
Összefoglalás
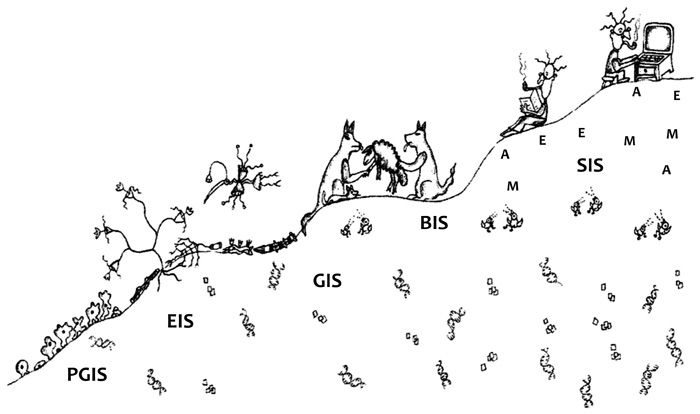
Az élővilág történetében és az evolúció nagy lépéseiben megváltozott
az élőlények térbeli szerveződése. Az élőlényeket mintázatok hálózatai
alkotják. A mintázatok legáltalánosabb értelemben rendezett időbeli
vagy térbeli inhomogenitások. Sejtekben molekuláris eloszlások,
soksejtűekben a sejtdifferenciáció térbeli szerveződése, állati
társadalmakban pedig a kasztok vagy a szociális aktivitás eredményei,
például a termeszvárak vagy hangyabolyok karakterisztikus mintákat
mutatnak. A minták átalakulásának okai öröklődési rendszerek hatásai,
fizikai kémiai, embrionális és ökológiai tényezők együttes változásai.
A mintázatok az organizáció egyik fő építőkövei. Ezért a minták és a
formák az élővilág sója és legkáprázatosabb
show-jai.
Az élővilág és a biológia rövid története
Az élővilág története sokféleképp mesélhető el. A mi történetünk
veleje röviden megfogalmazva az, hogy az élővilág története
mintázatok története. Az evolúció különböző stádiumaiban eltérő
mintázatok keletkeztek. Az élet történetének legmarkánsabb lépéseit
általában az öröklődés fő evolúciós lépéseire alapozzák (Maynard Smith
– Szathmáry, 1995; Jablonka – Lamb, 2005). Az öröklődés fő lépései
túlegyszerűsítve az 1. ábrán láthatók.
Az öröklődés megváltozásai azonban nem önmagukban,
elszigetelten mennek végbe. Eva Jablonka magyarázata csak egy
evolúciós tulajdonságra redukálja az evolúció magyarázatát a háromból:
az öröklődés megváltozására. Az öröklődés csak egy sajátosság az
evolúció három fő tulajdonságából: az öröklődésből, a változatosságból
és a szaporodásból. Csak az a rendszer képes evolválni, amely ezt a
hármasságot együtt mutatja. Mintázatok egyaránt felismerhetők
öröklődésben, változatosságban és szaporodásban (ivaros és ivartalan
szaporodás formájában). Változatok lehetnek molekulák, sejtek,
soksejtűek vagy társadalmak különbségei. Most itt a változatosság
keletkezésének megvilágítására összpontosítunk. A mintázatok ezekben a
változatokban megnyilatkozó rendet, szabályosságokat tükrözik,
inhomogenitások, szimmetriák, aszimmetriák, polaritás, kezesség,
topológia és szervezettség alakjában. Legáltalánosabban fogalmazva, a
mintázatok, rövidebben a minták, térbeli elrendezések (rendezett
inhomogenitások vagy heterogenitások). A pattern formation =
mintázatképződés fogalom hivatalos keresztapja Lewis Wolpert, angol
embriológus, aki 1969-ben vezette be ezt a fogalmat. A mintázatok
szerveződést vagy organizációt jelentenek, ami az élet forrása.
Most arról számolunk be, hogy a mintázatok
evolúciós átalakulása hogyan és miért ment végbe. Ahhoz, hogy ehhez a
történethez hozzáfoghassunk, röviden vissza kell nyúlnunk a biológia
történetének egyes kiemelkedő állomásaihoz. A biológia rövid története
nagyrészt híres képek, ábrák, diagrammok, modellek és metaforák
története. Igen híres, közismert képek D’Arcy Thompson parafinba
csöppentett olajból álló, élőkhöz hasonló műmedúzái. Hasonlóan
emlékezetes August Weismann csíra-szóma szétválást bemutató ábrája.
Méltán híres Conrad Hal Waddington epigenetikai tájképe, ami egy
hegyes tájból a völgyekbe legördülő golyót ábrázol. Ezzel Waddington
egy fejlődő élőlény történetét szemléltette, és ezen a mechanikai
allegórián keresztül az embriók előtt álló választási lehetőségeket
mutatta be. A hegyből legördülő golyó a fejlődési pálya (ontogenetikai
trajektória) előképe volt. Az evolúciót leggyakrabban a rátermettség
tájképén való mozgásként mutatják be, ami Sewall Wright leleménye. Ám
az egyik leghíresebb kép Ernst Haeckel nevéhez kapcsolódik. Ez a
gerinces embriók összehasonlító ábrázolása. A kép azt fejezi ki, hogy
az élőlények egyedi története, vagyis egyedfejlődése és evolúciója
(törzsfejődése) között grandiózus párhuzam van. A párhuzam abban áll,
vélte Haeckel, hogy az egyedfejlődés rövidített formában megismétli az
evolúció fő lépéseit. Ha aktualizálva összevetjük Haeckel képét a mai
tudásunkkal, a következőt láthatjuk:
A fejlődő embriók már egyedfejlődésük kezdetén is
eltérhetnek egymástól, akár közeli, akár távoli rokonságban álló
fajokba tartoznak. Az embriók különböznek megtermékenyített petéik
méretében és összetételében, aminek életmenetbeli és ökológiai okai
vannak. Az embriók fejlődésük egy meghatározott (ún. filotipikus)
stádiumában hasonlítanak leginkább egymáshoz (ez persze nem mindig van
így, legalább három modell írja le az embriók közti hasonlóság
eloszlását zigótától a kifejlett alakokig). Az embriók, fejlődésük
végeztével és a kifejlett állapot elérésével, egyre inkább eltérnek
egymástól.
Ezek a megfigyelések, az egyes szervezetek
történetének különbségei és hasonlóságai, magyarázatot igényelnek.
Ezek a magyarázatok ma még nem állnak rendelkezésünkre, de alapvetően
örökletes és ökológiai tényezők hasonlóságára és különbségére
próbáljuk ezeket visszavezetni. A biológia történetének leghíresebb
képeiben az a közös, hogy mintázatok leírását tartalmazzák.
Manapság közkeletű nézet, hogy az egyedfejlődés és
az evolúció genetikailag meghatározott folyamatok. A genetikát
általánosabban öröklődésnek kell mondanunk, mert nemcsak
nukleinsav-alapú átöröklés létezik, hanem ún. epigenetikai,
viselkedési és szimbolikus (például nyelvi) öröklődést is ismerünk
(Jablonka – Lamb, 2005). Ezek egymással kölcsönhatásban érvényesülnek.
Az öröklődésen alapuló magyarázatokkal szemben gyakran azt a kifogást
támasztják, hogy a gének nem kódolhatnak geometriai elrendezéseket,
formákat és mintázatokat. Ezért az öröklődésen alapuló magyarázatok
kiegészülnek fizikai kémiai mintázatképző okok és ökológiai tényezők
hatásaival.
A mintázatok természete
A mintázatok Wolpert megfogalmazásában a sejtdifferenciáció térbeli
szerveződését jelentik. Ez a meghatározás soksejtűekre vonatkozik. De
mintázatok az élettelen világban, az egysejtűekben, valamint
társadalmakban, kultúrákban és ökológiai rendszerekben is jelen vannak
és megfigyelhetők. Sokszor, mint például egy ezer pontból álló
ponthalmazban, még azt sem könnyű felismerni, hogy létezik-e
egyáltalán mintázat, vagy pusztán a képzeletünk játszik velünk.
Ilyenkor különféle tesztekhez folyamodhatunk, melyek alkalmasak
mintázatok kinyerésére a jelek özönéből. A legegyszerűbb
soksejtűekben, mint a Volvoxban, kétféle sejttípus van: testi és
szaporító sejtek. A Volvox képviseli a soksejtűség esszenciáját, ha
ilyesmi létezik. A többi soksejtű élőlény a volvoxos logika
kibővítésén, egyfajta dekorációján alapszik. Az ember négy
csíralemezében (ektodermában, endodermában, mezodermában, velősáncban)
411-féle sejttípus van, amelyből 145-féle idegsejt. Ezek között elvben
411 × 411 sejtes átmenet létezhet. A 411-féle sejt különféle
sejtvonalakon keresztül jön lére (a sejtvonal a sejtek leszármazásának
útvonala). A sejtvonal azonban önmagában még nem jelent mintázatot,
ehhez az egyes sejteknek megfelelő térbeli helyzetet kell
elfoglalniuk. Az evolúcióban az új sejtek vagy őssejtekből, vagy
differenciált sejtekből keletkezhetnek. Egy egyszerű mintázat
természetét mindenki képes önmagán is megérteni. Az emberi kezek és
lábak azonos sejttípusból állnak, de a kéz és a láb ugyanazokat a
sejteket más-más arányban és elhelyezkedésben tartalmazzák. A
gerincesek végtagjai (jelentős leegyszerűsítéssel élve) úgy
evolválnak, hogy megváltoznak a csontok építőkövei és azok kapcsolatai
képződésének időbeli (heterokróniás) viszonyai. A csontok fejlődése
kondenzálódó porcsejtekből indul el. A kondenzált porcszövet ezután
elágazásokat (bifurkációt) mutat, és a csontok részekre tagolódnak
(szegmentálódnak). A kondenzáció, az elágazás és a részekre tagolódás
időbeli variációi építik fel az uszonyok, a szárnyak és a lábak
változatosságát. A láb csontmintázata meghatározott alakot vesz fel,
és jellegzetes formát mutat. A formák képződésének megértéséhez az út
a minták megértésén át vezet.
A mintázatok változatossága
Az élő és élettelen természet nagyszámú mintázatból épül fel.
Mintázatokat alkotnak színek, a testek felszínei, a testek részekre
(például szelvényekre) tagolódása, és a testfüggelékek (például
pikkelyek, tollak, szőrök). Sokszor megfigyelhető, hogy a szervezetek
ismétlődő egységekből állnak. A szervezetek abban térnek el egymástól,
hogy más-más testrészek ismétlődnek, amelyek különböző megjelenésűek
(színük, méretük, alakjuk, elrendeződésük eltérő). Ilyen ismétlődő
egységek ízeltlábúakban és gerincesekben a testszelvények. Rovarokban
a testszelvények száma tíz és száz közt is változhat, míg gerinceseken
ez a szám kisebb. A szelvényekhez speciális testrészek kapcsolódnak,
mint rovarokban a szárnyak, csápok vagy lábak, gerincesekben pedig a
bordák. A mintázatok az evolúcióban azért fontosak, mert a változatok
fő alkotórészei.
Meglepő, hogy közös elemekből épülnek fel
komplikált, abiotikus szerkezetek, mint például a kémiai mintákban
szereplő szinguláris, pontszerű elemek, gömbök, sávok, csíkok, körök,
elágazó mintaelemek, spirálisok, hélixek, amelyek változatos
kombinációkat alkothatnak.
Mi lehet a fő motivációja a mintázatok
megismerésének? A biológia legtöbb problémája ún. terv-visszafejtéses
eljáráson alapszik. Ez azt jelenti, hogy egy megfigyelt mintázatból
megpróbáljuk megérteni a mintázatot előállító folyamatot. Ez nem
egyszerű eljárás, mert ugyanazt a mintázatot számos folyamat
előállíthatja. Így például a különböző lábak csontelrendeződései vagy
a bőrszervek (tollak, pikkelyek, szőrök, körmök, teknők, mirigyek)
hasonló mechanizmusokkal írhatók le. Általában az a kiindulópontunk,
hogy egy véletlenszerű elrendezést sztochasztikus folyamatok, míg az
ettől való eltéréseket szelekció állítja elő, evolúciós léptékben.
Ennek eldöntésére számos tesztet dolgoztak ki.
Mintázatok dinamikája
A mintázatokat mintázatképződési mechanizmusok állítják elő. Különböző
evolúciós stádiumokban és eltérő térskálákon más-más minták
keletkeztek. Elemi biológiai mintázatképződési
mechanizmusok a következők: reakció-diffúziós rendszerek,
előmintázatok, embrionális indukció, generikus fizikai kémiai
folyamatok, mint az ülepedés, konvekció, gravitáció, adhézió, fázis
szeparáció, és a pozicionális információs mechanizmus.
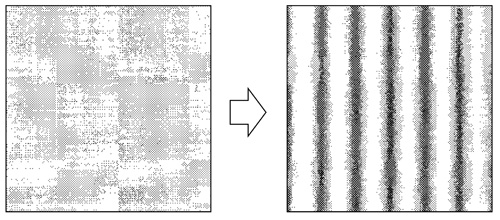
A mintázatképződés lényege, vagyis a mintázatképző
mechanizmusok hatása leegyszerűsíthető egy általános kijelentésre: a
mintázatképződés szimmetriasértésen alapul, amit a
2. ábra mutat be. Ez a helyzet egy sima
tükrű tó esetében. Ha ebbe kavicsot dobunk, onnan koncentrikus
körökből álló hullámok indulnak ki. A mintázatképződés megértéséhez
teljesen mindegy, hogy a tó tejből, vízből vagy alkoholból áll-e. A
mintázatképződés elsődleges kérdése az, hogy a sima, homogén,
szimmetrikus víztükrön szimmetriasértő instabilitások jönnek létre,
amelyek vízhullámokat keltenek. Fontos kérdés az is, hogy honnan
származik a tó vize vagy maga a kő, és hogy a vízfelszín egyáltalán
mennyire tekinthető eleve simának, homogénnek. A természet mintázatait
úgy teremti meg, hogy szimmetriasértő műveleteit folyamatosan
megismétli. Az sem egyszerű kérdés, hogy ezek a szimmetriasértő
mechanizmusok honnan erednek. Nem ismert teljes bizonyossággal
egyetlen biológiai mintázatképző mechanizmus evolúciós eredete sem.
Mintázatképződési változók a mintázatok
keletkezését kormányzó anyagok koncentrációja. Molekuláris szinten a
mintázatképződés a mintaképződésben szereplő ún. morfogén molekulák
reakciója és mozgása (diffúziója) kombinálódásából szerveződik. Ez a
lelke a Turing–Gierer–Meinhardt-hármas által leírt modellcsaládnak
(Meinhardt, 1982). Sejtek szintjén a fő mintázatképződési változók a
sejtek sűrűsége és mozgása.
A biológiai mintázatok eredetének
szemléleti keretei
A mintázatok evolúciós eredetének legalább hét felfogása alakult ki,
amelyeket itt röviden összefoglalunk.
1. Mintázatok születhetnek olyan képződményekből,
melyek maguk nem mintázatok, vagy keletkezhetnek már létező
mintázatokból.
2. Új minták új mintaképződési mechanizmusok
feltárásával állíthatók elő. Ez a kép árnyaltabbá tehető, ha
megfontoljuk az ősök és a leszármazottaik közti mintázatképződési
folyamatok kombinatorikáját. Ősi mintákból ősi minták jöhetnek létre
minták és generáló mechanizmusaik konzerválásával. Ősiből leszármazott
minták keletkezhetnek evolúciós újdonságok kialakulásával (ezt a korai
képet bővítjük itt most ki). Leszármazott mintákból ősi minták
jöhetnek létre ősökre való visszaütéssel vagy atavizmussal.
Leszármazott mintákból leszármazott minták képződhetnek mintázatok és
mechanizmusaik (pl. stabilizáló szelekciós) fenntartásával.
3. Egy másik nézet szerint a mintázatképződés
digitális→analóg állapotváltozásokon nyugszik. E felfogás hátterében
az a feltevés húzódik meg, hogy a mintaképződésben a DNS egydimenziós
információtartalma háromdimenziós szervezetekké fordítódik le. Azonban
a digitális-analóg kombinatorika ettől gazdagabb. Digitális→digitális
átmenetek is mintaképződéshez vezethetnek, például replikációkor.
Nemrégiben erre az alapra helyeződött a molekuláris evolúció egyik
irányzata. Analóg→analóg mintázatátmenetek alakulhatnak ki hullámok
egymásba történő átalakulásakor, például álló és utazó hullámok
konverziójában. Végül, analóg→ digitális átmenetek képződhetnek
például röntgensugaras mutagenezisben.
4. A negyedik lehetőség a mintázatok eredetének
leírására a következő: mintaképződéskor a sejtállapot (s) és a sejtek
helyzete (p) közt létesül kapcsolat. Ennek a kapcsolatnak három
formája lehet.
i. A helyzet megszabja a sejtek állapotát, ez a p→s
kapcsolat. Ilyenek az átrendeződéssel kapcsolatos mintázatképződési
módszerek, mint a differenciális sejtadhézió vagy Sperry
kemoaffinitási hipotézise.
ii. Az állapot megszabja a sejt helyzetét, ez az
s→p kapcsolat. Ilyen a pozicionális információ A pozicionális
információs mechanizmus illusztrálja a mintázatképződés napjainkban
uralkodó képét. Eszerint a sejtek a térbeli helyzetüknek, az
egyedfejlődés során befutott történetüknek és genetikai állapotuknak
megfelelő sorsot szereznek, melyet egymásnak jelzéseken át mondanak
el. Ebben a Wolpert által kidolgozott modellben a mintázatképződés
kétlépéses folyamat:
1. lépés • Van egy sejtparaméter, ez a pozicionális
érték. A pozicionális érték hozzárendelődik a sejt helyzetéhez a
fejlődő embrióban.
2. lépés • A sejtek genomjuktól és történetüktől
függően értelmezik a pozicionális értéket. A sejtek leolvasnak egy
kémiai miliőt, és beáll a sejtállapot. Az értelmezést követően más-más
módon
|
|
|
differenciálódnak. Ebben a keretben nincs kényszer,
vagyis korlátozás, ami megszabná azt a lehetséges tartományt, amivé az
egyes sejtek differenciálódhatnak, és így szinte minden minta
lehetséges. Ebben a keretben a mintázat azért változik, mert a
pozicionális érték értelmezése megváltozik (megváltozhat a sejtek
örökítőanyaga és a sejtek története).
iii. A sejtek állapota és helyzete egy harmadik
tényezőtől függ, ez a z→(s,p) kapcsolat. Ilyen például a sejtvonalas
mintaképződés.
5. Egy további nézet három elemre épül: a mintákat
nagyszámú ágens építi fel, ezek kombinációi gazdagok, melyekből a
szelekció válogatja ki az életképeseket.
6. Ezen a képen már csak pár módosítást végezve,
további mintázatképződési eredetre vonatkozó hipotézisek sorozata
gyártható. E kép kiegészíthető mutációval, rekombinációval és
sodródással történő mintaképződéssel. A mintázat működési képessége
vagy megbízhatósága lehet a mintázat megvalósulásának kritériuma.
7. A legelterjedtebb felfogás szerint a mintázatok
forrása aktiváló és gátló természetű mintaképző anyagok (morfogének)
autokatalitikus önaktiválása és oldalirányú vagy laterális gátlása. A
morfogének, az autokatalízis és a laterális gátlás evolúciós eredete
és e három tényező összekapcsolódásának születése ismeretlen.
A sokféle mintázat ma három osztályba sorolható:
ezek a pontmintázatok, csíkok, cellák és rácsok, valamint
származékaik, végül más, ráccsal nem lefedhető minták (például ún.
oszcillonok).
A mintázatképződés evolúciós aspektusai
Mi a kapcsolat az evolúció története és a mintázatképződés között? Az
evolúció során új mintázatok képződnek. Az evolúció egyes szakaszaiban
meghatározott, jellegzetes mintázatok uralkodtak.
Ezen a ponton megfogalmazzuk ennek a közleménynek a
legfontosabb tézisét. Az evolúció nagy lépéseiben új szimmetriasértő
mechanizmusok és ezek nyomán új mintázatok keletkeznek. Ilyen lépések
a független replikátorok összekapcsolódása kromoszómákká, a
prokarióta→eukarióta átmenet, az ivartalan klónok ivaros népességekké
történő átalakulása, az egysejtű→soksejtű átmenet, valamint a magányos
egyedek állati kolóniákká történő átalakulása (társas lények; hangyák,
darazsak, méhek, termeszek) (Maynard Smith – Szathmáry, 1995). A
főemlősök emberré válásakor megszülető ember s vele a nyelv eredete is
hordozhatja jellegzetes mintázatok képződését (ilyet leírtak az agy
evolúciójában), de az itt körvonalazott tézis ezen a ponton a
legkevésbé illusztrálható látványosan, míg más evolúciós átmenetben ez
csaknem nyilvánvaló. Most ezeknek eredünk a nyomába a sejtek,
soksejtűek és állati társadalmak evolúciójában megjelenő néhány
karakterisztikus mintázat bemutatásával. Szükséges és elégséges
hangsúlyozni, hogy sejtekben sejtekre jellemző, soksejtűekben
soksejtűekre (vagyis sejtcsoportokra) jellemző, társadalmakban pedig
társadalmakra jellemző mintázatok jelentek meg az evolúció nagy
lépéseiben, és hogy ezek nem létezhettek korábban. Vagyis minden új
ún. evolúciós egység karakterisztikus mintázatkészlettel, ennélfogva
pedig jellegzetes változatkészlettel rendelkezik. Most röviden
áttekintjük a mintázatoknak az evolúcióban bekövetkezett
megváltozásait.
Egysejtű mintázatok
A prokarióták alakját és jellegzetes fő mintázatait a sejtjeikben levő
molekulák eloszlása szabja meg. A prokarióták (eubaktériumok és
archebaktériumok) testalakját a sejtfal és a sejtplazma
ozmózisnyomásának kölcsönhatása szabályozza. A prokarióták alakvilága
a viszonylag merev sejtfal miatt eléggé szegényes, bár alakviláguk
gazdagsága nincs feltárva. Jellegzetes prokarióta mintázattal a
sejtfal rendelkezik, amit gyakran egy szőtteshez is hasonlítanak. A
molekuláris heterogenitás eloszlása is ismeretes. Például, ismert a
különböző vegyületeknek a bakteriális sejt ozmotikus nyomásához való
hozzájárulása. Eszerint a DNS, az RNS, a riboszómák, a tRNS, a
proteinek, az ionok, az oldatok ozmózishoz való hozzájárulása az
oldatok irányában növekvő mértékű. Prokarióták és eukarióták eltérő
ozmotikus stratégiákat mutatnak. Nemrégiben világossá vált, hogy a
prokarióták sejtpólusai indukcióhoz hasonló magatartást mutatnak, ami
segít a sejtosztódás megszervezésében. A sejt pólusai periodikus
minták kiindulópontjai lehetnek. A sejtek sejtciklusban élnek. Az
élővilág történetének öthatoda egysejtű állapotban telt el. Így az
élet történetének legnagyobb részét az egysejtűekre jellemző minták
uralták.
A protistákban a prokarióta-eukarióta sejtek
átmenete során kialakult a pókhálószerű sejtváz. A sejtváz, a
sejtfalak és a sejtmembránok kölcsönhatása rendkívül gazdag forma- és
mintázatvilágot hozott létre. A protisták lenyűgözően gazdag
sejtfelszíni és sejten belüli mintázatokkal rendelkeznek. Egysejtű
eukarióták uralkodó sejtmintázatai a membránok
kompartmentalizációjával keletkeznek. A kompartmentalizáció evolúciós
modellje lehet membrándinamikai természetű, ami leírja a membránok
hierarchikus begyűrődését a sejtek belső terébe. (Egy másféle
kompartmentalizációs modell a sejtek membránnal elhatárolt területeit
ún. paralóg expanzióval magyarázza meg). Meglepő egy híres sejtkutató
véleménye, miszerint a membrán kompartmentek száma nem változott az
eukarióta sejt evolúciós megjelenése óta. Ennek biztosan ellentmond a
különböző plasztiszok keletkezésével megjelenő további sejtes
térfelosztás. A protisták híresek sejtfelszíni mintázataikról: a
csillókról, ostorokról, az alapi testekről, a pigmentekről, a
pílusokról és más testfüggelékekről. A soksejtűekben a
mintázatképződést leggyakrabban a differenciális génkifejeződéssel
magyarázzák, amit rendszerint kiegészítenek különböző fizikai kémiai
mintázó mechanizmusokkal, mint a reakció-diffúzió, a konvekció, az
ülepedés, a fázisszeparáció, sejtadhézió és mások. A genetika és a
fizikai kémia együttesen segít megérteni, hogy miképp alakul ki
emberben a körülbelül 410-féle sejttípus térbeli elrendeződése. Ez a
differenciális génexpressziós magyarázat biztosan nem működik
egysejtűekben, mert egyetlen sejten belül ez megoldhatatlan. Sokan
keresik azokat az alapelveket, melyek közösek egysejtűekben és
soksejtűekben egyaránt. Egy ilyen közös alapelv lehet a pozicionális
információn alapuló mintázatképződés, egysejtűekben és soksejtűekben
egyaránt, valamint tipikus (generikus) fizikai kémiai
mintázatképződési mechanizmusok univerzális jelenléte.
Soksejtű mintázatok
Az egysejtűek után az evolúcióban megjelentek a soksejtűek. Soksejtűek
a gombák, a növények és az állatok. Ez az evolúciós átmenet legalább
tizennégyszer végbemehetett. A soksejtűek egyedfejlődésen mennek át.
Az egyedfejlődés egy pályát fut be, ez az egyedfejlődési pálya, amit
egyedfejlődési mechanizmusok hajtanak zigótától a kifejlett
szervezetig (feltéve, hogy létezik ilyen kifejlett állapot, ami inkább
csak unitér szervezetekben jellegzetes (ettől a moduláris szervezetek
különböznek). A soksejtűek szintjén megfogalmazható mintaképződés a
sejtosztódásból, a sejtkölcsönhatásokból és a sejthalálból
kovácsolódik össze. Ezek a komponensek ősibb, korábbi mintaképződési
módokat bővítettek ki a soksejtűek sejtcsoportjai megszületésének
pillanatában.
Ha az egyes soksejtűek élete az egyedfejlődési
pályákban zajlik, akkor a soksejtűek evolúciója pályamódosításnak
tekinthető. Ilyen pályamódosító mechanizmusok az allometria
(differenciális testméretnövekedés különböző térbeli irányokba), a
heterokrónia (az egyedfejlődési események kezdőpontjainak,
végpontjainak és rátáinak megváltozása), az embrionális indukció, a
homeózis (egyes testrészek más testrésszé, például lábnak csáppá
történő átalakítása, a jelátviteli változások, a génszabályozási
megváltozások, a génkifejeződési régiók eltolódása a testtengelyek
mentén, vagy a gének számának növekedése, valamint a transzkripciós
faktor variánsok átalakulása, mint a homeotikus, Sox, POU, T-box
génekben és géncsaládokban.
A gombák testalakját a fal és a fal által határolt
anyag kölcsönhatása szabja meg. A fal képlékenysége szabja meg, hogy
hol lehet alakváltozás a gombák testében. A fal képlékenységét gének
irányítják, de ma még nem tudjuk, hogyan. A fal képlékenységén át a
testalak szabályozása nem csupán gombasajátosság vagy specialitás.
Hasonló elven nyugszik mindazon lények alakjának szabályozása, amelyek
fallal rendelkeznek, vagyis prokarióták, sok protista, gomba s növény
egyaránt.
A növények testalakját is a merev fal és a
sejtjeikben levő folyadék uralja. Ha az egyes sejteket egy-egy
képlékeny, de merev falú gömbnek tekintjük, akkor a növények egy
metafórája a gömbhálózat lehetne (ezt másképp csatolt hidrosztátnak
vagy sejtes szilárd anyagoknak – cellular solids – is nevezik).
Az állatok epitéliumból álló hólyagcsírából
fejlődnek ki, melyek gasztruláció alatt test alaptervet és abban
szervkezdeményeket fejlesztenek ki. Az állatok sokfélesége embrióik,
és a belőlük fejlődő egyedfejlődési pályáik gazdagságán alapszik. Az
egyedfejődési pályákon az embriók fejlődését egyedfejlődési
mechanizmusok hajtják. Ilyen mechanizmusok a sejtadhézió, a
sejtjelzések, az indukció, a különböző, ún. generikus fizikai kémiai
mechanizmusok, mint a fázisszétválás, a reakció-diffúzió, az ülepedés,
a gravitáció és az anyagkeveredés.
Milyen fő mintázatképződési
mechanizmusok működnek a soksejtűekben, és ezeken belül az állatok
evolúciójában (mert ez ismert), és ezeknek milyen a rendszertani
csoportok közti eloszlása? Az állatok mintázatképződésének vázlatos
áttekintése meggyőzően illusztrálja a soksejtűkre jellemző,
karakterisztikus mintázatokat és mintázatképződési mechanizmusokat,
melyek egysejtűekben még nem létezhettek.
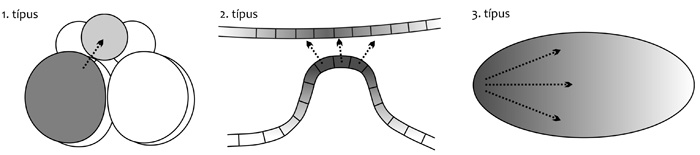
Egy átfogó kép szerint az állati embriogenezisnek
három fő módszere alakult ki, melyeknek a rendszertani eloszlása is
ismert (Davidson, 1991) (4. ábra). A
három fő embriogenezismód a következő. Az 1. típusú embriogenezisben
az osztódó megtermékenyített petesejtből alapító sejtek képződnek
(invariáns barázdálódással). Az alapító sejtek sejtvonalakat hoznak
létre. A sejtvonalak a sejt leszármazási útvonalai. A sejtvonalak
fontos szerepet játszanak a korai embriók térbeli szerveződésében. A
sejtek differenciációja önállóan és feltételes módon történik,
többnyire sejtvándorlás nélkül. Feltételes esetben a
sejtdifferenciáció sejtpárok közti (indukciós) jelátvitellel megy
végbe. Az ilyen embriók mozaikosak, azaz az embriók részei viszonylag
függetlenül fejlődnek egymástól. Az embriók kb. tíz sejtosztódás után
jönnek létre. Az ilyen embriók főképp gerinctelenekre jellemzők, de
minden állati törzsben előfordulnak, bár rovarokban és gerincesekben
ritkábbak. A 2. típusú embriogenezis sejtcsoportok közt megy végbe
(változatos barázdálódással), helyi induktív kölcsönhatásokkal és
sejtek közti jelátvitellel, ez jellemző a gerincesekre. A 3. típusú
embriogenezist transzkripciós faktorok és más
mintaképző molekulák diffúziója kormányozza, és változatos
szincíciumokban megy végbe (amelyben a sok sejtmag még nem határolódik
el sejtmembránokkal egymástól), és jellegzetes képviselőik a rovarok.
A 2. és 3. embriogenezis típus a 1.-ből alakulhatott ki a mintázódás
felgyorsításával. A 3. csoportban a mintázódás oly gyors, hogy sok
élőlény még nem is soksejtű, máris megkezdődik a testszelvények
határainak lefektetése. Az egyedfejlődés és az evolúció kapcsolatának
egy mai képét az 5. ábrán foglaltuk össze.
Társadalmi mintázatok
A soksejtűek közül az állatok csoportokat, kolóniákat és társadalmakat
alkothatnak. A társadalmak mintázatait legegyszerűbb a rovarok
kasztjaiban megérteni. A kasztok képződéséhez a testarányok
megváltozása vezet, ami egyaránt áll genetikai és környezeti
szabályozás alatt. Az egyes kasztokba tartozó élőlények arányai
alapvető jelentőségűek a munkamegosztásban. A munkamegosztás során az
egyes kolóniatagok magatartásaikat ún. kompozit struktúrákba
szervezik, melyek soros, párhuzamos vagy ezekből képződő vegyes
kapcsolású elrendezések. A kompozitok a kasztképződés hatékonyságát
jelentősen befolyásolják. Párhuzamosan elvégzett munkák megnövelik a
kolóniák hatékonyságát. A kompozitok univerzálisan előforduló
szerkezetek, melyek szinte minden térbeli skálán megfigyelhetők
(Molnár, 2001).
Jellegzetes mintázatokat alkotnak a társas élet
szereplői szociális kölcsönhatásaik megvalósításában. A társas életet
is, mint szinte minden evolúciós átmenetet, az együttműködés és a
konfliktusok együttélése jellemzi. Az ilyen helyzetek kezelésére
alakult ki az együttműködés és a csalásban testet öltő konfliktusok
stratégiai leírása. A társas magatartási formák négy fő kategóriája, a
kölcsönös előnyösség, az önzetlenség, az önzés és a rosszindulat,
valamint ezek sorozatai, a szociális csoportok egyik legfőbb vonása.
Jellegzetesen társas minta a szaporodási aránytorzulás. Ez akkor
keletkezik, ha bizonyos élőlények nagyobb arányban járulnak hozzá az
utódok génjeihez, mint mások. Ugyancsak karakterisztikus társas minta
a szociogenezis, ami alatt egy kolónia növekedését és fejlődését
értjük. Kolóniák képződésekor szuperorganizmus fejlődik ki.
Szuperorganizmusban új fajta munkamegosztás (szociális anatómia) és új
típusú, csoportszintű koordináció (szociális élettan) születik meg,
egyedek hálózataiból. Társadalmak szintjén lépnek fel a kolóniákon
belüli és a kolóniák közötti kölcsönhatások, például táplálékelosztás,
védelmek és támadások.
Állati társadalmakban, kiváltképp pedig emberben,
létrejött a kultúra. A kultúra több hagyomány együttese, mely szintén
bővelkedik mintázatokban. Ilyenek az oktatás, az életmód, az étkezés,
az öltözködés, az utazás, és az ember által készített művek,
műtermékek és alkotások sokasága.
Kulcsszavak: mintázatképződés, evolúciós átmenetek,
szimmetriasértés, embriók evolúciója, genetikai tool-kit, evo devo
IRODALOM
Carroll, Sean B. (2005): Endless Forms
Most Beautiful. The New Science of Evo Devo. Norton, New York
Davidson, Eric H. (1991) Spatial
Mechanisms of Gene Regulation in Metazoan Embryos. Development. 113,
1–26.
Jablonka, Eva – Lamb, Marion. (2005):
Evolution in Four Dimensions. MIT Press, Cambridge
Maynard Smith, John – Szathmáry Eörs.
(1995): The Major Transitions in Evolution. WH Freeman, Oxford
Meinhardt, Hans. (1982): Models of
Biological Pattern Formation. Academic Press, London
Molnár, István (2001) The Reliability
Theoretical Aspects of Evolution. Nagy és Társa, Eger
|
|