|
genetikai összetételére vezethető vissza. A
dendrogramok ugyanakkor egyértelműen azt tükrözik, hogy az erdélyi
populációk elkülönülése sokkal nagyobb mértékű a lápi-, mint a díszes
tarkalepke esetében.
Eredményeinket összefoglalva elmondhatjuk tehát,
hogy a genetikai variabilitás szintje a két fajban nem mutatja
egyértelműen a fenotípusos plaszticitásukban, valamint a habitat- és a
tápnövényigényükben megnyilvánuló különbségek következményeit.
Ugyanakkor a differenciálódásuk mintázata világosan tükrözi a
sztochasztikus hatások erősségében meglévő különbségeket.
3. esettanulmány: A nagy tarkalepke (Melitaea phoebe) és a
Kovács-tarkalepke (M. telona kovacsi) genetikai és morfometriai
varianciájának szerkezete
A nagy tarkalepke széles elterjedésű faj, amely áreáján belül számos
alfajra tagolódik (Pelz, 1995; Wahlberg – Zimmermann, 2000). Hazánkban
két nemzedéke van, nyáron a hernyók folyamatosan táplálkoznak, majd
bebábozódnak, és az új nemzedék már nyár derekán megjelenik.
Fenotípusos plaszticitása viszonylag nagy, aminek következtében
különböző zavart élőhelyeken, akár gyomos legelőkön, útszegélyeken is
előfordul. Tápnövényválasztása széleskörű, hernyói számos aszat
(Cirsium spp.) és imola (Centaurea ssp.) fajjal
táplálkoznak. Egy-egy élőhelyen többnyire nem nagy egyedsűrűségű, de
nagy diszperziós képessége miatt előfordulása egyenletesnek mondható.
A Kovács-tarkalepke taxonómiai helyzete hosszú időn keresztül vitatott
volt, önálló faji státusza a 2005–2006 között megjelent közlemények
alapján vált elfogadottá (Varga et al., 2005; Russel et al., 2006). A
Kovács-tarkalepke egynemzedékű faj, hernyója folyamatosan átnyaral és
áttelel. A Kovács-tarkalepke hazai élőhelyein monofág, hernyói
gyakorlatilag kizárólag a magyar aszattal (Cirsium pannonicum)
táplálkoznak. Előfordulására jellemző, hogy lokálisan, foltokban
jelenik meg, ahol populációit rendszerint nagy egyedsűrűség jellemzi.
Diszperziós képességét limitálja, hogy erősen tápnövényhez kötött
(Tóth et al.. 2010, in print). A két faj diszperziós képességét,
tápnövény- és élőhelyigényét tekintve tehát jelentősen különbözik
egymástól. Arra számítottunk, hogy ezeknek a különbségeknek a
következményei a fajok variabilitásának szerkezetében is
megnyilvánulnak. Az alacsonyabb szintű génáramlás következtében a
Kovács-tarkalepke populációi között erősebb az izoláció mértéke.
Emiatt bennük alacsonyabb szintű variabilitást, közöttük viszont
intenzívebb differenciálódást vártunk, mint a nagy tarkalepke
esetében. Ugyanakkor a nagyobb elterjedés következtében feltételeztük,
hogy a nagy tarkalepke populációkban kifejezettebb a szubpopulációs
tagolódás, mint a lokálisan előforduló
Kovács-tarkalepke-populációkban.
Mindkét faj mintáit az Aggteleki-karszt régióban
gyűjtöttük. A genetikai és morfometriai vizsgálatokban is két
Kovács-tarkalepke és három nagy tarkalepke populáció mintáit
vizsgáltuk meg. Tekintettel a Kovács-tarkalepke faji státusának
korábbi bizonytalanságára, elsődleges célunk az volt, hogy a két faj
közötti genetikai és morfometriai differenciálódást megvizsgáljuk.
Ezen túlmenően azonban teszteltük kezdeti hipotézisünket is.
A genetikai variabilitás szintjének összehasonlító
elemzése egyértelműen alátámasztotta feltételezésünket. A fajok
allélkészletét tekintve a nagy tarkalepke populációkban jelentősen
több allélt (tizenöt lókuszban ötvenegy) detektáltunk, mint a
Kovács-tarkalepkében (ugyanezen lókuszokban harminckettő), annak
ellenére, hogy ez utóbbi fajnál a mintavételek három egymást követő
évben megismétlődtek, míg a nagy tarkalepke három populációjában csak
egy-egy évben gyűjtöttünk mintát. Az allélok frekvencia szerinti
megoszlása a két fajban szintén világos eltéréseket mutatott. A
Kovács-tarkalepke populációkban viszonylag sok (28,8%) fixálódott
allélt találtunk, míg a nagy tarkalepke populációkban kisebb volt az
arányuk (12,4%). A genetikai variancia szerkezetének tanulmányozása
során összehasonlítottuk a Wright-féle F-statisztika indexeit.
Eredményeink arra utaltak, hogy a nagy tarkalepke populációkban
jelentős heterozigóta-hiány volt, ami nem volt jellemző a
Kovács-tarkalepkére. A heterozigóta-hiány számos evolúciós hatás
következménye lehet. Ezek egyike az, hogy a
populációk nem homogének, hanem szubpopulációs tagolódást mutatnak. A
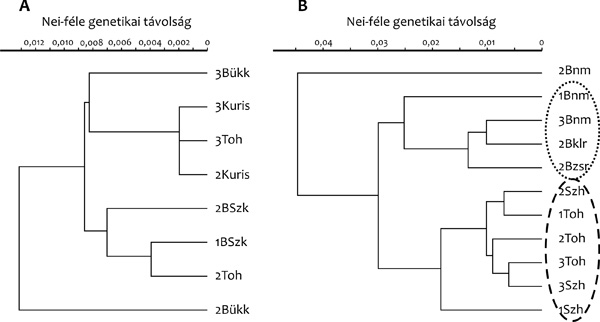
Nei-féle genetikai távolságok mátrixából szerkesztett dendrogram a két
faj egyértelmű elkülönülését mutatta (3.A ábra).
Ugyanakkor a két fajon belül a dendrogram nem mutatott jelentős
különbséget a differenciálódás mértékében.
A morfometriai vizsgálatok eredményei a genetikai
eredményeket sok tekintetben alátámasztották. Bár statisztikai
szempontból egyértelmű különbséget nem tudtunk kimutatni a két faj
között, bizonyos tendenciákat mégis észleltünk. A vizsgált tizenhat
jellegből nyolc több mint 10%-kal variábilisabbnak mutatkozott a nagy
tarkalepke populációkban, mint a Kovács-tarkalepkében. A fenotípusos
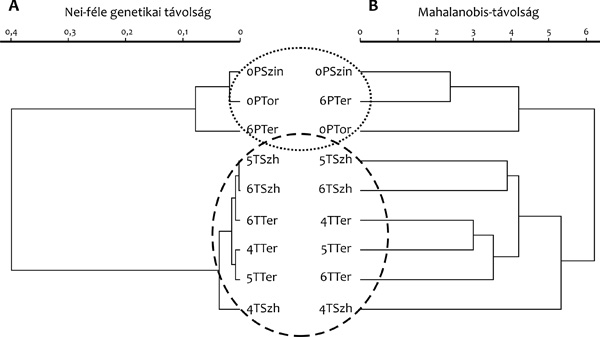
variancia szerkezetét a Mahalanobis-távolságok alapján szerkesztett
fenogramokon tanulmányoztuk. A genetikai adatok alapján szerkesztett
dendrogramhoz hasonlóan a fenogramokon is egyértelmű elkülönülés
mutatkozott a két faj között (3.B ábra).
Eredményeink alapján megállapíthatjuk, hogy a
korábbi taxonómiai és ökológiai vizsgálatokkal összhangban mind a
morfometriai, mind pedig a genetikai adatok alátámasztják a
Kovács-tarkalepke faji státusát. A két faj variabilitásának szintjére
megfogalmazott hipotézisünket a genetikai és a morfometriai
vizsgálatok eredményei egyaránt alátámasztják. A heterozigóta-hiány
mértékében mutatkozó különbségek pedig a két faj
populációstruktúrájában meglévő eltérésekre utalnak. Ugyanakkor a
vizsgált populációk kevés száma nem tette lehetővé, hogy a
differenciálódás földrajzi mintázatában feltételezett különbségeket is
igazolni tudjuk.
4. Tanulságok és további lehetőségek
Mint várható volt, vizsgálataink nagyrészt igazolták előzetes
elvárásainkat. Ezért általánosítható eredményeink alapján, ha
mutatkoznak ilyenek, el kell gondolkodnunk azon, hogyan léphetünk
innen tovább. Ilyen lehetőségeknek az alábbiak kínálkozhatnak.
Láttuk, hogy a szürkés hangyaboglárka és a bükki
szerecsenboglárka esetében lényegében ugyanaz a területi elkülönülés
mutatkozik a bükk-hegységi és az aggteleki-karszti populációk között.
Változatosságukban és a területenkénti genetikai differenciálódás
mértékében azonban lényeges különbségek mutatkoztak. Ezeket jórészt
megmagyarázhattuk az életmenetükben tapasztalható jelentős
eltérésekkel: az obligát, illetve fakultatív mirmekofiliával. Ebből az
életmenet-különbségből azonban az is adódik, hogy a bükk-fennsíki
rétek kezeletlenség okozta degradációjára, a jelek szerint, a kettős
függőségű (tápnövény + hangyagazda) hangyaboglárka-populáció
érzékenyebben reagált, mint a kevésbé tápnövényspecialista és csak
kevéssé hangyafüggő szerecsenboglárka. Láthattuk tehát, hogy a két
esetben a nagyjából ugyanolyan mértékű área-szétszakadozás eltérő
mintázatot generál a populációk szerkezetében és genetikai
változatosságában. Az élőhelykezelési prioritások meghatározásában
tehát célszerű az érzékenyebb indikátorfaj alapján elindulni.
A lápi tarkalepkénél végzett eddigi genetikai
vizsgálataink alapján legalábbis erősen gyanítható, hogy a
Kárpát-medence legutóbbi jégkorszak utáni benépesedése nem egyetlen
gócterületből ment végbe. Feltűnő egyrészt, hogy ez a faj a medence
nyugati-délnyugati részén a szinte síkvidéki-völgyi helyzetű (Dráva,
Mura, Zala stb.) területektől egészen a magaskarsztokig (Magas-Bakony,
Szlovéniai-karszt) és a szubalpin-öv rétjeiig (Stájerország)
eredetileg, legalábbis potenciálisan szinte összefüggő nagy
területeket népesíthetett be, ugyanakkor a „középdunai flóraválasztót”
nemhogy nem lépi át sehol, de meg sem közelíti. Mint ahogy, másrészt,
keleti irányból sem lépi át az Erdélyi-szigethegységet mint hasonló
rangú faunaválasztót. Ez az eredmény legalább két dologgal nagyon
erősen egybecseng. Elterjedéstörténeti szempontból azzal a ténnyel,
amit már több, természetvédelmi szempontból is lényeges fajnál
sikerült igazolni, hogy a Kárpát-medence posztglaciálisan,
genetikailag is jól igazolhatóan, több önálló gócból népesült be, mint
ahogy ezt például a kis apollólepkénél (Parnassius mnemosyne) és a
kerekfoltú szerecsenlepkénél (Erebia medusa), gerincesek közül a
mocsári békánál (Rana arvalis), az erdei pocoknál (Chlethrionomys
rutilus) láthattuk. Evolúciógenetikai szempontból pedig ismételten
aláhúzza a növényvilágunk és faunánk diverzitásában is kulcsfontosságú
„preillír-prealpin” területeink önálló evolúciós-differenciálódási
gócterület értékét. Amiből természetvédelmünknek máris le kell vonnia
azokat a gyakorlati következtetéseket, amelyek szerint nem egyszerűen
„fajmegőrzési programokban” kell gondolkodnia és cselekednie, hanem
máris hatékonyan kell törekednie a hagyományos taxonómiai szemlélet
szerinti „fajokon” belüli evolúciósan jelentős populációcsoportok, az
ún. evolúciósan szignifikáns egységek (ESU) megőrzésére.
A nagy és a Kovács-tarkalepkénél vizsgálataink
mindenekelőtt arra világítottak rá, hogy az átfedő külső- és
genitália-morfológiai jellegek ellenére közöttük szinte bizonyos a
génáramlás teljes hiánya. Ez egyrészt a két faj közti ökológiai
(tápnövény, életciklus, populációstruktúra) eltérések részletesebb
elemzésére irányíthatja figyelmünket, amelyen jelenleg is dolgozunk.
Másrészt viszont további hipotéziseket is megfogalmazhatunk arra
vonatkozóan, hogy melyek lehetnek azok a szaporodásbiológiai és
genetikai tényezők, amelyek mintegy „ráerősítenek” a koegzisztáló
populációk ökológiai elkülönülésére.
Kulcsszavak: szürkés hangyaboglárka, bükki szerecsenboglárka, lápi
és díszes tarkalepke, nagy és Kovács-tarkalepke, enzim polimorfizmus
IRODALOM
Benes, Jiri et al. (eds.) (2002):
Butterflies of the Czech Republic: Distribution And Conservation.
I–II. SOM, Prague
Cizek, Oldrich – Konvicka, Martin (2005): What is a Patch in a Dynamic
Metapopulation? Mobility of an Endangered Woodland Butterfly,
Euphydryas maturna. Ecography. 28, 791–800.
Elmes, Graham W. & Thomas, Jeremy A.
(1987): Die Biologie und Ökologie der Ameisen der Gattung Myrmica. In:
Schweizerische Bund für Naturschutz: Tagfalter und ihre Lebensräume:
Arten, Gefährdung, Schutz. Basel, 404–9.
Elmes, Graham W. – Thomas, J. A. –
Wardlaw, J. C. (1991a): Larvae of Maculinea rebeli, a Large Blue
Butterfly and Their Myrmica Host Ants. patterns of Growth and
Survival. Journal of Zoology (London). 224, 79–92.
Elmes, Graham W. – Wardlaw, J. C. –
Thomas, J. A. (1991b): Larvae of Maculinea rebeli, a Large Blue
Butterfly and Their Myrmica Host Ants. Wild Adoption and Behaviour in
Ants Nests. Journal of Zoology (London). 223, 447–460.
Elmes, Graham W. – Thomas, J. A. –
Hammarstedt, O. – Munguira, M. L. – Martin, J. – van der Made, J.
(1994): Differences in Host Ant Specificity between Spanish, Dutch and
Swedish Populations of the Endangered Butterfly, Maculinea alcon (Den.
& Schiff., 1775) (Lepidoptera). Memorabilia zoologica. 48, 55–98.
Fowles, Adrian P. – Smith, Richard G.
(2006): Mapping the Habitat Quality of Patch Networks for the Marsh
Fritillary Euphydryas aurinia (Rottemburg, 1775) (Lepidoptera,
Nymphalidae) in Wales. Journal of Insect Conservation. 10, 161–177.
van Halder, Inge – Barbaro, L. – Corcket,
E. – Jactel, H. (2008): Importance of Semi-natural Habitats for the
Conservation of Butterfly Communities in Landscapes Dominated by Pine
Plantations. Biodiversity and Conservation. 17, 1149–1169.
Konvicka, Martin – Hula, V. – Fric Z.
(2003): Habitat of Prehibernating Larvae of the Endangered Butterfly
Euphydryas aurinia (Lepidoptera: Nymphalidae): What Can Be Learned
from Vegetation Composition and Architecture? European Journal of
Entomology. 100, 313–322.
Konvička, Martin – Čížek, O. – Filipová,
L. – Fric, J. – Beneš, J. – Křupka, M. – Zámečník, J. – Dočkalová, Z.
(2005): For Whom the Bells Toll: Demography of the Last Population of
the Butterfly Euphydryas maturna in the Czech Rep. Biologia. 60, 5,
1–7.
Liu, W. H. – Wang, Y. F. –Xu, R. M.
(2006): Habitat Utilization by Ovipositing Females and Larvae of the
Marsh fritillary (Euphydryas aurinia) in a Mosaic of Meadows and
Croplands. Journal of Insect Conservation. 10, 351–360.
Malicky, Hans (1969): Übersicht über
Präimaginalstadien, Bionomie und Ökologie der mitteleuropäischen
Lycaenidae (Lepidoptera). Mitteilungen der entomologischen
Gesellschaft Basel. 19, 25–91.
Pelz, Volker (1995): Biosystematik der
europäischen Arten der Tribus Melitaeini Newman, 1870. Oedippus
(Gesellschaft für Schmetterlingsschutz, Bad Neustadt). 11, 1–62.
Russell, Peter – Tennent, W. J. – Pateman,
J. – Varga, Z. – Benyamini, D. – Pe’er, G. – Bálint, Zs. –
Gascoigne-Pees, M. (2007): Further Investigations into Melitaea
telona Fruhstorfer, 1908 (= ogygia Fruhstorfer, 1908; = emipunica
Verity, 1919 (Lepidoptera: Nymphalidae), with Observations on Biology
and Distribution. Entomologist’s Gazette. 58, 137–166.
Thomas, Jeremy A. – Wardlaw, Judith C.
(1992): The Capacity of a Myrmica Ant Nest to Support a Predacious
Species of Maculinea Butterfly. Oecologia. 91, 101–109.
Varga Zoltán – Szabó S. & Kozma P. (2005):
Melitaea ogygia kovacsi Varga 1967 (Lepidoptera: Nymphalidae) in the
Pannonian Region: Taxonomy, Binomy, Conservation Biology. In: Kühn, E.
– Feldmann, R. – Thomas, J. – Settele, J. (eds.): Studies on the
Ecology and Conservation of Butterflies in Europe. Vol. I: General
Concepts and Case Studies. Conference Proceedings UFZ Leipzig-Halle,
December 2005. 65–68.
Wahlberg, Niklas – Zimmermann, Marie
(2000): Pattern of Phylogenetic Relationships Among Members of the
Tribe Melitaeini (Lepidoptera: Nymphalidae) Inferred from
Mitochondrial DNA Sequences. Cladistics. 16, 347–363.
Weidemann, Hans-Josef (1986): Tagfalter 1. Entwicklung-Lebensweise.
Neumann-Neudamm, Melsungen
|