|
|
A nyelv eredetének problémája
Az emberi nyelv az élővilágban egyedülálló jelenség, melynek
kétségkívül vannak biológiai alapjai. Az emberszabású majmok, delfinek
vagy papagájok több évi alapos képzés alatt sem képesek olyan szinten
elsajátítani, mint amilyet az embergyerekek spontán magukévá tesznek.
Néhányan úgy vélik, hogy intelligenciánk önmagában is elég nyelvi
képességünk kialakulásához, ám nem magyarázzák meg azt, hogy ez az
intelligencia miben különbözik például az emberszabású majmokétól.
Szerintünk inkább az okozza a különbségeket, hogy az embernek olyan
neuronális kapacitásai vannak, melyek a többi állatfajban csak nagyon
kezdetleges formában vannak jelen. Ennek következtében többek között
hatékonyan tudunk kezelni hierarchikus struktúrákat: ilyenek a
szintaxis rekurzív elemei a nyelvben, az alegység-stratégia az
eszközkészítés során és a másod-, harmad- és negyedrendű
intencionalitás az elmeteóriában.
A nyelv olyan összetett rendszer, amely
párhuzamosan koordinál több képességet is, ezért a nyelv genetikai
eredetének kiderítéséhez sok más érzékelési, mozgató- és kognitív
rendszert kell megismernünk; azt, hogy ezek hogyan változtak
külön-külön és hogyan változott a közöttük levő kapcsolat. A nyelv
eredetének vizsgálatát azonban éppen ezen ismeretek hiánya
akadályozza. A nyelvészek között sincs általános egyetértés arról,
hogyan írják le a nyelvet – különféle megközelítések léteznek, melyek
képviselői heves csatározásokat vívnak. Mi, biológusok, úgy tartjuk,
hogy a legkisebb közös nevező ebben a vitában az lehet, ha a nyelvet
mint a szimbolikus referencia és az összetett szintaxis kombinációját
határozzuk meg.
A nyelv létrejöttének több olyan előfeltétele is
van, melyek a jelen pillanatban nem különösebben érdekesek számunkra –
például az emberszabású majmoknak nincs lesüllyedt gégefőjük, emiatt
nem tudnak olyan sokféle fonémát képezni, és a vokalizációjuk sincs
teljesen agykérgi kontroll alatt. Az ember evolúciója során
kétségtelenül kialakultak ezek a tulajdonságok, ám ezek nem
feltétlenül szükségesek a nyelvhez, mint olyanhoz. Tökéletesen
működőképes nyelvet lehetne létrehozni kevesebb fonémából vagy akár
vokalizáció nélkül is (például jelnyelv). Minket elsősorban a nyelvi
operátorok neuronális implementációja érdekel, modalitástól
függetlenül. Nehéz elképzelni a nyelv eredetét a tanítás, az utánzás
képessége és fejlett elmeteória nélkül. Az emberszabású majmok ezen
képességei korlátozottak, az emberfélék evolúciója során pedig
valószínűleg éppen azért mentek keresztül jelentős változásokon, mert
a nyelvvel együtt evolválódtak. Ehhez még hozzáadódik egy fontos
emberi adaptáció, a nagy, nem rokon csoportokban való kooperáció
képessége (Maynard Smith – Szathmáry, 1995). Ezek a tulajdonságok
együtt egy olyan adaptív csomagot alkotnak, mely csak az emberre
jellemző, és melynek megléte valószínűbbé teszi a nyelv kialakulását
(Szathmáry – Számadó, 2008).
Ami a nyelv neurobiológiai alapjait illeti, néhány
tankönyv még mindig azt a túlegyszerűsített nézetet képviseli,
miszerint az agy bal féltekéjében található Wernicke-mező a
szemantikáért, a Broca-mező pedig a szintaxisért felelős. Valójában a
nyelvi összetevők agyi lokalizációja nagyon is rugalmas: a teljes bal
agyfélteke eltávolítása pár hónapos korban meglepően kevés elváltozást
okoz a nyelvi képességekben, mivel a jobb félteke képes átvenni a
szükséges feladatokat. A nyelv komponensei az egyedfejlődés során
lokalizálódnak az agyban, méghozzá a rendelkezésre álló
„legkényelmesebb” helyen (vö. Karmiloff-Smith, 2006). A nyelv, mint
egy amőba, megtalálja az agyban a számára legmegfelelőbb élőhelyet. Az
emberi agynak egy elég nagy része alkalmas élőhely a „nyelvi amőba”
számára, a főemlősök agyában viszont – úgy tűnik – nincs helye
(Szathmáry, 2001).
A nyelv eredete máig megoldatlan probléma: azon
kívül, hogy igen kevéssé támaszkodhatunk a fosszíliákra, a genetikai
és fiziológiai kísérletezés is erősen korlátozott embereken és
emberszabású majmokon. Ezen kívül a nyelv egyedülálló volta
lehetetlenné teszi a szigorú értelemben vett összehasonlító
tanulmányokat, melyek a biológia más területein oly hasznosnak
bizonyulnak. Éppen ezért van kiemelt szerepe a nyelv eredetének
kutatásában a szimulációknak: az elméleti kutató feladata az, hogy
megpróbálja modellezni a nyelv előnyelvből való kialakulásának
közbülső lépéseit. Ebben a cikkben a nyelv problémakörének genetikai,
neurobiológiai és evolúciós hátterét szeretnénk áttekinteni.
A gének és a nyelv
A humán és a csimpánz genom szekvenálása befejeződött, ám rengeteg
munka van még hátra. A gének ismerete még nem minden, azt is ki kell
deríteni, hogy a fenotípus hogyan feleltethető meg a genotípusnak. A
gének kölcsönhatásban vannak: kifejeződésüket nem csak a környezet,
hanem egyéb gének is befolyásolják, így a kölcsönhatások hálózata
szintén rendkívül fontos. A kérdés az, hogy a gének hatása hogyan
szivárog fölfelé, egészen addig, míg valamilyen tulajdonságban,
viselkedésben megnyilvánulnak.
A nyelvi képesség olyan tulajdonság, ami minden
emberben megvan, és többnyire csak kvantitatív különbségeket lehet
felfedezni az egyes személyek között. A kérdés, hogy ezért mennyiben
felelősek a gének, és mennyiben a környezet. Erre a kérdésre részben
ikervizsgálatokból kaphatunk választ. A nyelvi rendellenességek egy
része egypetéjű ikrekben többnyire együtt jelenik meg, míg kétpetéjű
ikrekben nem, ami azt jelenti, hogy a géneknek nagyobb hatásuk van egy
rendellenesség létrejöttére/hiányára, mint a környezetnek. Ilyen
például a diszlexia és a specifikus nyelvi zavar (Specific Language
Impairment – SLI). Egészséges ikrekkel végzett kísérletekből pedig
kiderült, hogy a fonológiai rövid távú memóriában, artikulációban,
szókincsben és morfoszintaxisban észlelt variancia mögött is részben
genetikai hatások állnak.
Bizonyára voltak genetikai változások az emberfélék
evolúciója során, melyek fellendítették a nyelv evolúcióját, de
általános, aspecifikus módon, tehát általában nem konkrét agyterületek
konkrét jellegzetességeire hatva. Ilyenek lehettek az agyméretet
befolyásoló gének, vagy egy nemrég felfedezett gén, mely talán
közreműködik az emberi agykéreg hatrétegű szerkezetének kialakításában
– hozzátesszük, önmagában egyik sem lehet felelős a nyelv
megjelenéséért.
Tudunk azonban olyan változásokról is, melyek
közvetlenül hatnak a nyelvre. Myrna Gopnik 1990-ben azonosította a
FOXP2 gént, melynek egy domináns mutációja ún. fejlődési verbális
dyspraxiát (Developmental Verbal Dyspraxia – DVD) okozott egy angol
anyanyelvű családban (Gopnik, 1990). Ez nemcsak a száj- és arcizmok
renyheségét okozta, hanem a családtagok nyelvhasználatát is
befolyásolta: az érintett személyek nem voltak képesek olyan általános
szabályokat automatizált formában elsajátítani, mint hogy az igék múlt
idejét az -ed, a főnevek többesszámát pedig az -s végződéssel
képezzék. Ehelyett minden egyes esetet külön kellett memorizálniuk,
úgy, mint az egészséges embereknek a rendhagyó ragozású szavakat.
A FOXP2 gén terméke minden gerinces állatban
megvan, és bizonyított, hogy pozitív szelekción ment keresztül az
elmúlt kétszázezer évben. Más fajok vizsgálatából kiderült, hogy a
FoxP2 gén a szenzomotoros integrációért és a mozgás irányításáért
felelős idegsejthálózatok fejlődéséért felelős, ami arra enged
következtetni, hogy a nyelv eredetének a mozgásirányításhoz is köze
van. Énekesmadarakban a FoxP2 fontos szerepet játszik egyes
agyterületek fejlődésében és szezonális aktivitásában. További
érdekesség, hogy az énekesmadarak énektanulásáért felelős idegdúc
analógnak tűnik az ember bazális ganglionjával, mely a verbális
tanulásban játszik szerepet. Mind a FoxP2, mind az azonos géncsaládba
tartozó FoxP1 gén az ember és az énekesmadár agynak funkcionálisan
hasonló agyterületein fejeződik ki, melyek a szenzomotoros
integrációért és a mozgásirányításért felelősek. Mindezeken túl a
FoxP2 gén kiesése madarakban pontatlan és hiányos vokális imitációt,
mutációja egerekben az ultrahangos vokalizáció rendellenességét
okozza, melyet a kisegerek az anyjuktól való szeparációkor használnak.
A génnek tehát más fajokban is a szociális kommunikációhoz van köze. A
gén terméke olyan egyéb géneket szabályoz, melyek a szinaptikus
plaszticitásban, az idegsejtek fejlődésében, az idegsejtnyúlványok
irányításában és az ingerületátadásban játszanak szerepet.
A nyelvi képesség biológiai motivációjú felosztása
nagyon fontos lenne ahhoz, hogy meg tudjuk határozni, mik lehettek a
nyelv közbülső fejlődési állapotai. A nyelvészeti elméleteket a nyelv
biológiájával összefüggésben kéne kialakítani. Sajnos eddig nem
született olyan elmélet, mely részletesen figyelembe vette volna az
agyi mechanizmusokat.
Az agy és a nyelv
Az idegsejtek aktivitásának vizsgálata rohamos fejlődésen ment
keresztül az utóbbi évtizedekben. A műszerek pontossága folyamatosan
nő, és egyre gyakrabban használják őket arra, hogy megállapítsák, mely
agyterületek aktívak különböző nyelvi műveletek végzése közben. Bár a
különböző agyi funkciók különböző agyi területek aktivitásával
függenek össze, fontos kiemelni, hogy ez nem jelenti a funkciók
genetikailag szigorúan kódolt lokalizációját. Az agy fejlődése – főleg
az élet első néhány hónapjában – nagyon plasztikus, így sok kognitív
képesség szinte teljesen helyre tud állni korai agysérülések után.
Ugyanez igaz a nyelv különböző összetevőire, így a szintaxisra is. Még
sérülések hiányában is a nyelv feldolgozása szétosztva, az agy több
területén történik, az agykérgen kívül az agykéreg alatti területek is
szerepet játszanak ebben, például a bazális ganglion és a kisagy.
Elképzelhető, hogy az eszközkészítésnek is szerepe
volt a nyelv evolúciójában (Greenfield, 1991), tehát a hatékony
eszközkészítés irányába ható szelekció egyben elősegítette a nyelv
evolúcióját is és vice versa. Érdekes, hogy amikor három különböző
méretű poharat kell egymásba rakniuk, az emberszabású majmok és a
kisgyerekek ezt mindig az ún. „fazékmódszerrel” oldják meg, egyszerre
csak egy poharat mozgatva: először a legnagyobba teszik a középsőt,
majd ebbe a legkisebb poharat. A gyerekek viszont egy idő után
elsajátítják az ún. „alegység-módszert” is: először a legkisebb
poharat teszik a középsőbe, majd a kettőt egyszerre mozgatva teszik a
legnagyobba. Ez utóbbi stratégia rekurzívnak tekinthető, melynél a
cselekvés „nyelvtana” hasonló szerkezetű, mint a rekurzív szabályokkal
szerkesztett mondataink. Tecumseh W. Fitch és Marc D. Hauser (2004)
kísérlete szerint rekurzív szabálynak megfelelő mintázatokat
mesterséges nyelvben tamarin majmok nem voltak képesek felismerni, az
emberek viszont igen. E két jelenség arra utal, hogy a majmok agyában
még nincsenek meg azok a más idegsejteket szabályozó neurális
kapcsolatok, melyek lehetővé tennék a rekurzív műveletek elvégzését.
Egy másik kísérletben megkérték a kísérleti
alanyokat, hogy készítsenek eszközöket a 2,5 millió éves olduvai
kultúrára jellemző módszerekkel, és közben azt figyelték, hogy mely
agyterületeik aktiválódnak. Érdekes módon nemcsak a premotor cortex
aktiválódott, hanem a Broca-mező hátsó része is, ami tovább erősíti
azt az elképzelést, hogy van kapcsolat az eszközkészítés és a nyelv
között.
Az agy és a nyelv koevolúciója
Be kell látnunk: nem értjük, pontosan hogyan is működik az agy.
Fejlődését mindenképp befolyásolják genetikai tényezők, de így is
nagyon plasztikus: még az egypetéjű ikrek agya is meglepően
különbözik. Jean-Pierre Changeux (1983) szerint a felnőtt agykéreg
funkcionális mikroanatómiája a kezdeti szinapszistúltengésnek, majd a
felesleges szinapszisok és idegsejtek funkcionális kritériumokon
alapuló szelektív eliminációjának eredménye (tehát a nemhasznált
szinapszisok törlődnek).
Az eddigiekből kiderült, hogy az emberi agynak
nagyon nagy része képes nyelvi információkat, sőt nyelvtani
operációkat is feldolgozni. Ez azt jelenti, hogy nincs egy olyan előre
rögzített makroanatómiai struktúra, ami kizárólagosan végezné a
nyelvvel kapcsolatos feladatokat. Sokkal inkább arról lehet szó, hogy
a nyelv feldolgozásának képessége valamilyen funkcionális
mikroanatómiai struktúrától függ, az idegsejtek kapcsolatainak
valamely statisztikai-hálózati tulajdonságától, mely az agy nagy
részére jellemző (Szathmáry, 2001). Hogy ez a többi állat agyához
képest új tulajdonság minek az eredménye, nem tudjuk. Szelekcionista
szemszögből három lehetőség van: már az idegsejtek kezdeti kapcsolatai
is újszerűek; az idegsejtek kezdeti kapcsolatai nem újszerűek az
állatokéhoz képest, de a szelekciós mechanizmus új; illetve mind a
kezdeti kapcsolatok, mind a szelekciós mechanizmus új.
|
|
|
A fenti elképzelés szoros összefüggésben van az agy
és kogníció koevolúciójának újabb elméletével. A tradicionális
elképzelés szerint a koevolúció „lentről felfelé” mechanizmus alapján
zajlik: eszerint ha egy idegi struktúra genetikai változáson megy
keresztül, akkor kognitív teljesítménye alapján a természetes
szelekció vagy lehetővé teszi az elterjedését, vagy nem. A „felülről
lefelé” mechanizmus azonban Stanley I. Rapoport szerint sokkal nagyobb
szerepet játszott az emberi kognitív képességek, így a nyelv
evolúciójában is: eszerint, mivel az agy egyedfejlődése plasztikus, az
olyan területein, melyek nagyobb igénybevételnek vannak kitéve,
kevesebb idegsejt, illetve idegsejtkapcsolat fog leépülni, ami jobb
teljesítményhez vezet. Ilyen körülmények között bármilyen genetikai
változás, mely az adott agyterület növekedéséhez vezet, kedvező.
Ez az elmélet gyakorlatilag egy korábbi elmélet
kiterjesztése, miszerint a komplex teljesítményekre képes agy
megváltoztatja a szelektív környezetet (mivel az állat környezetét
többnyire a hasonló aggyal rendelkező fajtársai alkotják), ami aztán
még összetettebb agyra fog szelektálni. Tehát a szelekció fő
hajtóereje nem a fizikai, hanem a szociális környezet lesz, vagyis a
fajtársak viselkedése. Másrészt ez a mechanizmus a genetikai
asszimilációnak is nagyon szép példája, miszerint „a tanulás vezeti az
evolúciót”. Ez azt jelenti, hogy egyes újítások, melyeket eleinte
minden egyednek meg kellett tanulnia, később genetikailag kódolódnak
az agyban. Mivel azonban a nyelvi rendszerek nagyon gyorsan változnak,
kicsi a valószínűsége, hogy a genom asszimilálni tudjon konkrét
nyelvtani szabályokat. Itt inkább egy általános feldolgozási
mechanizmus genetikai asszimilációjára kell gondolni, ami a szabályok
pontosabb feldolgozására, gyorsabb megtanulására tette képessé az
egyedeket (Szathmáry, 2001).
Az eddigieket összefoglalva javaslataink tehát a
következők:
• A nyelv kialakulásához genetikai változásokra
volt szükség, melyek befolyásolták az agy jelentős részének
egyedfejlődését.
• Ezek a változások az itt található idegsejtek
kapcsolatainak statisztikai mintázatát, valamint dinamikus fejlődését
érintették.
• Az agy egyedfejlődésének szelekciós plaszticitása
következtében a nyelv és az agy koevolúciója a nyelvtani szabályok
feldolgozása képességének genetikai asszimilációjához vezetett.
A nyelv és az evolúció
Az ember adaptív csomagja
A nyelven és a komplex eszközhasználaton kívül kizárólagos emberi
tulajdonság a hatékony tanítás, az imitáció, a fejlett
elmeteória és a nagyméretű, nem rokoni csoportokon belüli
kooperáció képessége is (Maynard Smith – Szathmáry, 1995). Véleményünk
szerint ezek a tulajdonságok nem véletlenül jelentek meg együttesen.
Adaptív csomagot alkotnak, mely valószínűleg szinergisztikus módon,
koevolúcióval fejlődött az elmúlt ötmillió évben (Szathmáry – Számadó,
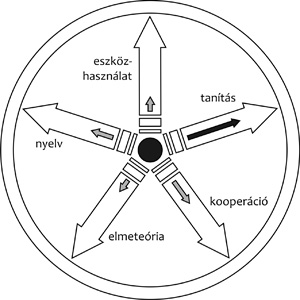
2008). Az 1. ábra „koevolúciós kereke”
azt szimbolizálja, hogy bármelyik „küllő” (tulajdonság) evolúciója
előnyére válhatott a többi tulajdonságnak is, még akkor is, ha a
szelekció többször küllőt váltott, felváltva fókuszálva az egyes
tulajdonságokra. Így nincs is sok értelme azt a kérdést boncolgatni,
hogy milyen sorrendben alakultak ki a küllők. Azt, hogy ezek a
tulajdonságok összekapcsolódnak, alátámasztja az autizmus esete: az
autizmussal élőknek az elmeteória, a kommunikáció és a nyelv területén
vannak hiányosságaik. Az a predikciónk, hogy több olyan gént is fogunk
találni a közeljövőben, amely pleiotróp hatású, tehát a koevolúciós
keréknek egyszerre több küllőjére is hat.
Az emberszabású majmok az emberrel ellentétben
nagyon kevéssé értik a fizikai világ ok-okozati összefüggéseit, és ez
többek között az eszközkészítési képességeiket is limitálja. Ezzel
szemben az ember már jobban érti a kauzális viszonyokat, és
lehetséges, hogy éppen ez tette lehetővé számára a nyelv
elsajátítását. Derek C. Penn és Daniel J. Povinelli (2007, 111.)
szerint sok új, és közvetlenül nem megfigyelhető kauzális viszonyt már
ismert jelenségek analógiáján keresztül értünk meg: például az atom
szerkezetét a naprendszerhez hasonlítjuk, az elektromosságot az áramló
folyadékhoz, a gravitációt a fizikai erőhöz.
Sok nyelvtani szerkezet is kauzálisan működik: az egyik elem
megjelenése egy másik elem megjelenését teszi kötelezővé. Másrészt a
nyelv is analóg szerkezetek végtelen variációján alapszik: Jancsi
szereti Juliskát, a cica szereti a tejet stb. Tehát a rekurzión kívül
az analóg érvelés is összeköti a nyelvet az eszközkészítéssel. További
fontos emberi képesség, hogy közös szándékokat vagyunk képesek
koordinálni, ami lehetővé teszi a kooperációt és a tanítást. Fontos
még a pontos mozgáskoordináció, ami többek között az eszközkészítéshez
szükséges.
Az emberre jellemző adaptív csomag elemeit két
csoportra lehet osztani: a nélkülözhetetlen procedurális komponensekre
(hierarchikus szerkezetek kezelésének képessége, analóg érvelés,
imitáció, közös szándékok és pontos mozgáskoordináció), és a komplex
adaptív képességekre (tanulékonyság, komplex kooperáció és elmeteória,
nyelv és eszközkészítés). Véleményünk szerint a kölcsönhatásoknak és
folyamatoknak ez az összetett hálózata pleiotróp hatású gének
közreműködésével alakulhatott ki. Egy ilyen gén, mely kedvező hatását
több tulajdonságra is kifejti, gyorsabban elterjed a populációban, és
rögzülése is valószínűbb, mint egy olyan génnek, mely csak egy
tulajdonságra hat.
Szelekciós forgatókönyvek
a nyelv kialakulására
Az emberi nyelv kialakulását számos elmélet magyarázza. A legtöbb
elmélet a nyelv funkcionális szerepét többé-kevésbé jól indokolja, így
ezen az alapon nem lehet eldönteni, hogy melyik elmélet a hasznosabb.
Ezt a problémát egy új játékelméleti kutatás segítségével szeretnénk
feloldani, mely figyelembe veszi a kommunikáló felek érdekeit és a
kommunikációs jelek költségét is.
Eszerint a kutatás szerint ha a jelek mindkét
félnek egyformán költségmentesek, akkor az őszinte jeladás csak akkor
lehet evolúciósan stabil, ha nincs érdekellentét a kommunikáló felek
között. Amint megjelenik az érdekellentét, a kommunikáló felek – mivel
ezt költségmentesen tehetik – csalni fognak (például egy harc előtti
kommunikáció során erősebbnek mutatják magukat, mint amilyenek a
valóságban, s így próbálják elriasztani az ellenfelet). Viszont ha a
jelek költsége a jeladó minőségétől függ, elképzelhető olyan eset,
hogy a költségmentes jelek őszinték maradnak, még akkor is, ha
érdekellentétről van szó (Számadó, 1999). Az emberi nyelv esetében a
legegyszerűbb ilyen eset az, amikor a nem őszinte jeladót megbüntetik,
amihez természetesen fel is kell ismerni a csalókat. Összegezve tehát,
megegyezésen alapuló, költségmentes jelek akkor maradhatnak fent, ha a
kommunikáló felek közös érdekekről vagy ütköző érdekek ellenőrizhető
vonatkozásairól kommunikálnak; egyéb esetekben a jelek költségesek
lesznek.
Mit mond nekünk ez az elmélet az emberi nyelv
megjelenéséről? A beszéd vagy a mutogatással való kommunikáció szinte
költségmentes, így azok az elméletek lehetnek helyesek, melyek szerint
az emberi nyelv megjelenésének kontextusában nem volt érdekellentét,
vagy melyek szerint volt ellentét, de a jelek őszinteségét könnyen
ellenőrizni lehetett. Az irodalomban található tizenegy legnépszerűbb
elméletet megvizsgálva, egyik sem felelt meg a második esetnek,
viszont több olyan elmélet is van, mely szerint nem volt érdekellentét
a kommunikáló felek között. Számadó Szabolcs és Szathmáry Eörs (2006)
három másik kritérium szerint tovább szűkítették a szóba jöhető
elméletek körét. Mivel nem valószínű, hogy az első szavak absztrakt
fogalmak lettek volna, a nyelv elsődleges céljának a fizikai
valóságra, a kommunikáló feleket körülvevő környezetre való utalásnak
kellett lennie. A harmadik szempont azt vizsgálja, hogy az elmélet
megmagyarázza-e az emberi nyelv általánosító képességét, tehát azt,
hogy gyakorlatilag bármiről tudunk beszélgetni, míg az állatok
többnyire csak saját állapotukról közölnek információt. A negyedik
kritérium szerint pedig az elméletnek meg kell tudnia magyarázni az
emberi nyelv egyedülállóságát az állatvilágban.
A legtöbb elmélet több kritériumnak sem felelt meg.
Számadó és Szathmáry (2006) csak két olyan elméletet találtak, amely
három kritériumnak is megfelelt: az egyik szerint az emberi nyelv az
eszközkészítés (Greenfield, 1991), a másik szerint a vadászat
(Washburn – Lancaster, 1968) kontextusában alakult ki. Ám egyik
elmélet sem magyarázza az emberi nyelv egyedülállóságát az
állatvilágban.
Sajnos egyik elméletet sem támasztották alá
modellezéssel, pedig a nyelv evolúciójának kutatásában ennek
kiemelkedő szerepet kellene játszania. Az elméletek igazolásához
mesterséges világokban kéne rekonstruálnunk a nyelv megjelenését. A
robotok használatának több előnye is lenne: a megjelenő szavak a
fizikai valósághoz kapcsolódhatnának, és a robotok szenzorok
segítségével érzékelhetnék cselekedeteik következményeit (Nolfi –
Floreano, 2002).
Az evolúció néhány nagy átmenete többször is
végbemehetett (például többsejtű szervezetek kialakulása vagy társas
életmód kialakulása az állatok között), míg mások egyszeriek. Ilyennek
tartjuk a nyelv megjelenését is (Maynard Smith – Szathmáry, 1995),
bár, mivel nem ismerünk minden ma élő és már kihalt élőlényt, a nyelv
kialakulásának egyszeriségét csak feltételezni tudjuk. Egy evolúciós
átmenet valószínűségét két tényező csökkentheti: a
variáció-limitáltság és a szelekció-limitáltság. Az előbbi azt
jelenti, hogy az átmenethez szükséges genetikai változások
valószínűsége kicsi, így a szelekció számára rendelkezésre álló
genetikai variánsok száma is kicsi. Az utóbbi esetben genetikai
variánsokban nincs hiány, viszont különleges környezet szükséges az
átmenetet biztosító változat fixálódásához. Az emberi nyelv
evolúciójának kutatásakor mindkét szempontot figyelembe kell vennünk:
milyen genetikai háttere van a nyelvnek, milyen variánsok lehettek a
szelekció alapanyagai a nyelv kialakulásakor, illetve milyen
körülmények vezettek ahhoz, hogy azok a variánsok maradtak fenn,
melyek lehetővé teszik, hogy a ma ismert nyelveken kommunikáljunk.
Kulcsszavak: nyelv, szintaxis, evolúció, FOXP2, plaszticitás, adaptív
csomag, eszközkészítés, vadászat
IRODALOM
Changeux, Jean-Pierre (1983) : L’homme
neuronal. Librairie Arthème Fayard, Paris
Fitch, W. Tecumseh – Hauser, Marc D.
(2004): Computational Constraints on Syntactic Processing in a
Nonhuman Primate. Science. 303, 377–380.
Gopnik, Myrna (1990): Feature-blind
Grammar and Dysphasia. Nature. 344, 715.
Greenfield, Patricia M. (1991): Language,
Tools and Brain: The Ontogeny and Phylogeny of Hierarchically
Organized Sequential Behaviour. Behavioral and Brain Sciences. 14,
531–595.
Karmiloff-Smith, Annette (2006): The
Tortuous Route from Genes to Behavior: A Neuroconstructivist Approach.
Cognitive, Affective, & Behavioral Neuroscience. 6, 9–17.
Maynard Smith, John – Szathmáry Eörs
(1995): The Major Transitions in Evolution. Freeman, Oxford
Nolfi, Stefano – Floreano, Dario (2002):
Synthesis of Autonomous Robots through Evolution. Trends in Cognitive
Sciences. 6, 31–37.
Penn, Derek C. – Povinelli, Daniel J.
(2007): Causal Cognition in Human and Nonhuman Animals: A Comparative,
Critical Review. Annual Review of Psychology. 58, 97–118.
Számadó Szabolcs – Szathmáry Eörs (2006):
Selective scenarios for the emergence of natural language. Trends in
Ecology and Evolution. 21, 555–561.
Számadó Szabolcs (1999): The Validity of
the Handicap Principle in Discrete Action-Response Games. Journal of
Theoretical Biology. 198, 593–602.
Szathmáry Eörs – Számadó Szabolcs (2008):
A Social History of Worlds. Nature. 456, 2–3.
Szathmáry Eörs (2001): Origin of the Human
Language Faculty: The Language Amoeba Hypothesis. In: Trabant, Jürgen
– Ward, Sean (eds.): New Essays on the Origin of Language. Mouton/de
Gruyter, New York, 41–51.
WEBCÍM >
Washburn, Sherwood Larned –
Lancaster, C. (1968): The Evolution of Hunting. In: DeVore, Irven –
Lee Richard B. (eds.): Man the Hunter. Aldine, Chicago, 293–303.
LÁBJEGYZET
* A cikk a következő
könyvfejezet alapján készült (jelentősen rövidítve): Fedor Anna –
Ittzés Péter – Szathmáry Eörs (2008): The Biological Background of
Syntax Evolution. In: Bickerton, Derek – Szathmáry Eörs (eds.):
Biological Foundations and Origin of Syntax. The MIT Press, Cambridge,
MA.
<
|
|