|
funkcióra történő specializálódása
(differenciálódása) kapcsán bekövetkező, a teljes kromatinra
kiterjedő, mélyreható kromatinszerkezeti változás is, mely a
nukleoszómákon belül a különböző hisztonok kohéziójának mértékét is
érintik (Meshorer et al., 2006).
Emlősök sejtjeiben a gének az őket vezérlő
DNS-szekvenciák metilációja által (bizonyos fehérjék kötőhelyéül
szolgálva) elhallgattathatók, így a metilációban részt vevő enzimeknek
az egész kromatinra kiterjedő szabályozó hatásuk van. A hisztonfarkak
modifikációi is a gének jelentős hányadát érinthetik. Az egyes
módosításokat végrehajtó enzimek hibás – például túlzott vagy
szabályozatlan – működése, az őket kódoló gének egyes daganatos
folyamatokra jellemző megváltozása miatt, hozzájárul a patológiás
helyzet kialakulásához és fenntartásához. A génátírást szabályozó
fehérjék általános (nem bázissorrend-specifikus) DNS-kötő
tulajdonsággal is rendelkeznek. Az ilyen aspecifikus kötőhelyek száma
és hozzáférhetősége a globális kromatinszerkezet függvényében
módosulhat, ami a specifikus kötőhelyekre kötődni képes faktorok
számát befolyásolhatja (Misteli, 2008).
A citoplazmatikus és magi kompartment közötti
kapcsolatok révén szintén a teljes kromatinállományt potenciálisan
érintő hatások érvényesülhetnek. A magban a makromolekulák óriási –
kb. 100 mg/ml-es – koncentrációban vannak jelen. Ennek
kísérletes befolyásolása egyes magi funkciók reverzibilis változását
vonja maga után, ami arra utal, hogy a magmembránon át folyó
transzportfolyamatok révén ennek a tényezőnek is fontos, globális
szabályozó szerepe lehet (Hancock – Hadj-Sahraoui, 2009). A
magmembránban számos, a sejthártyára jellemző ioncsatorna,
membránkötött enzim és jelátvivő zsíroldékony kismolekula található,
ami a sejthártyán keresztül zajló jelátvitellel analóg folyamatok magi
szerepére utalhat (Bustamante, 2006).8
Egyelőre nehezen ítélhető meg, hogy milyen globális vagy génspecifikus
szabályozási jelentősége lehet a citoplazma váz elemei és a mag
dinamikus belső váza között a magmembránon átívelő fehérje-fehérje
asszociációkon keresztül megvalósuló kapcsolatnak.
Globális változások hátterében akár egyetlen
géntermék domináns szerepe érhető tetten. A kromatint a sejtosztódás
céljaira mikroszkópban jól felismerhető kromoszómákba szervező
fehérjék egyikének genetikai hibái például súlyos és komplex fejlődési
rendellenességeket eredményeznek (Gard et al., 2009). A magon belüli
topológiai viszonyok meghatározásában önszerveződési elv látszik
érvényesülni, melynek eredményeként előálló rendkívül dinamikus
struktúra vázként is funkcionál (ezt szokás magmátrixnak nevezni
[Gluch et al., 2008]); ennek alkotásában a magmembránt bélelő hálózat,
a lamina is részt vesz, melynek génszabályozásban betöltött általános
szerepe abban is megmutatkozik, hogy az őt alkotó fehérjék hibás
szerkezetét okozó genetikai rendellenességek, mutációk igen komplex
következményekkel járnak (például a korai öregedés-szindrómák egyes
formáiban [Hale et al., 2008]).
Az előbbi jelenségek többféle optikán keresztül is
tanulmányozhatók: vizsgálhatók egy adott gén aktivációs
állapotát meghatározó molekuláris szereplők a
DNS-szekvenciaelemek, melyek részt vesznek az adott hatás
kiváltásában; a fehérjék, melyek a kromatinterület szerkezetét
meghatározzák; az egyes molekulák közötti kölcsönhatások stb. A
magasabb rendű szerveződés törvényszerűségeit azonban egyelőre nem
ismerjük kellőképpen ahhoz, hogy az adatokat ebben az összefüggésben
értelmezni tudjuk.
Az egyetlen metafora felé9
A természettudományok művelői egy-egy újabb jelentős, a területükön
tett felismerésről tudomást szerezve azt egyfajta evidenciaélményként
élik meg, és ezzel összefüggésben úgy is tűnik, mintha belátható
közelségbe kerülne a kérdéskör lényegi megoldása. Mégis egyre merőben
váratlan felfedezések születnek; vagyis paradox módon a „részek”
(egyes laborok) előrébb tartanak, mint a kollektív egész (a kutatói
közösség). Ugyanakkor a kollektív munka egyre összehangzóbb világkép
felé vezet, a hatalmas puzzle megfejtett részeivel egy, a bevált
természettudományos fogalmakkal és módszerekkel megfelelően leírható
történet körvonalai rajzolódnak ki.
A különböző szerveződési szinteken érvényesülő hatások és változások
egymáshoz való viszonyában fogható meg a magi szabályozási folyamatok
eddig talán legkevésbé feltárt és legizgalmasabb aspektusa. Az
eszköztár e viszonylatok megismerésére valószínűleg már rendelkezésre
áll. A magasabb rendű kromatinszerkezetet például közvetlenül
láthatóvá és kvantitatíve is jellemezhetővé teszik a különböző
kromoszómális régiók fizikai kapcsolatát (kémiai
keresztköthetőségükön, illetve térbeli közelségükön,
ko-lokalizációjukon keresztül) kimutatni képes molekuláris biológiai
és mikroszkópos technikák. A DNS-hez kötődő fehérjék „világítóvá”
tétele az élő sejten belül (fluoreszcens fehérjét kódoló
DNS-szakasszal megtoldott génjük bevitele által) a különböző
kölcsönhatásokban részt vevő fehérjék szerepéről ad – szó szerint –
felvilágosítást. Segítségükkel a genomikai technikák nyújtotta adatok
topológiai összefüggésbe helyezhetők, megérthető lesz az egyes
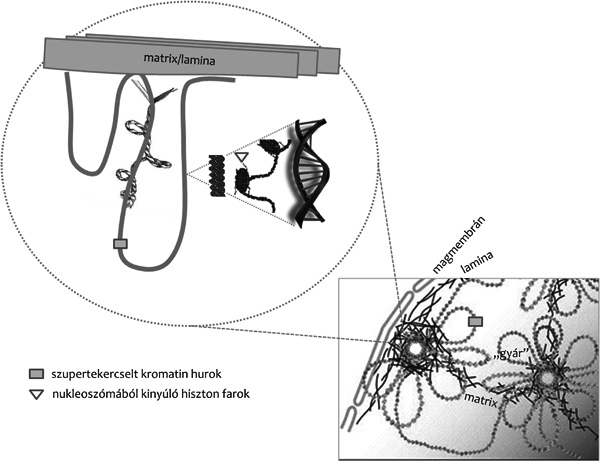
szerveződési szinteken (1. ábra) érvényesülő lokális és globális
hatások egymásra épülése, megtudjuk, hogy miként szerveződnek azok a
struktúrák, melyhez a különböző funkciókra szerveződő kromatinhurkok
kihorgonyzódnak, ez hogyan valósul meg és szabályozódik, miként
alakulnak ki a különböző feladatokra specializálódott
fehérjeegyüttesek, „gyárak”, hogyan vezérlődik és realizálódik ezek
kapcsolata a kromatin megfelelő elemeivel stb. A biokémiai,
molekuláris biológiai, genetikai kutatások keretében felfedezett
molekuláris szereplők és mechanizmusok is egyre gyarapítják azt a
„muníciót”, mellyel ezek az összefüggések és kérdések célba vehetők.
Ebben a megismerési folyamatban részt venni szellemi kaland,
örömforrás és kihívás, mely alapvető megfigyelésekkel kecsegtet
sokféle kutatói indíttatású és stílusú megközelítés számára.
Kulcsszavak: kromatinszerkezet, sejtmag, génszabályozás
IRODALOM
Bustamante, José Omar (2006): Current
Concepts in Nuclear Pore Electrophysiology. Canadian Journal of
Physiology and Pharmacology. 84, 3–4, 347–65.
WEBCÍM
>
Cook, Peter R. (2010): A Model for All
Genomes: The Role of Transcription Factories. J. of Molecular Biol.
395, 1, 1–10.
WEBCÍM
>
Dundr, Miroslav – Ospina, J. K. – Sung, M.
H. – John, Sam – Upender, M. – Ried, T. – Hager, G. L. – Matera, A. G.
(2007): Actin-Dependent Intranuclear Repositioning of an Active Gene
Locus in Vivo. Journal of Cell Biology. 179, 6,1095–103.
Gard, Scarlett – Light, W. – Xiong, B. –
Bose, T. – McNairn, A. J. – Harris, B. – Fleharty, B. – Seidel, C. –
Brickner, J. H. – Gerton, J. L. (2009): Cohesinopathy Mutations
Disrupt the Subnuclearorganization of Chromatin. Journal of Cell
Biology 187, 4, 455–62.
Gluch, Angela – Vidakovic, M. – Bode, J.
(2008): Scaffold/Matrix Attachment Regions (S/Mars): Relevance for
Disease and Therapy. Handbook of Experimental Pharmacology. 186,
67–103. By Diffusion
WEBCÍM >
Groth, Anja – Rocha, W. – Verreault, A. –
Almouzni, G. (2007): Chromatin Challenges During DNA Replication and
Repair. Cell. 128, 4, 721–33.
WEBCÍM >
Hager, Gordon L. – McNally, J.G. –
Misteli, T. (2009): Transcription Dynamics. Molecular Cell. 35, 6,
741–53. WEBCÍM
>
Hale, Christopher M. – Shrestha, A. L. –
Khatau, S. B. – Stewart-Hutchinson, P. J. – Hernandez, L. – Stewart,
C. L. – Hodzic, D. – Wirtz, D. (2008): Dysfunctional Connections
Between the Nucleus and the Actin and Microtubule Networks in
Laminopathic Models. Biophysical Journal. 95, 11, 5462–5475.
WEBCÍM
>
Hancock, Ronald – Hadj-Sahraoui, Yasmina
(2009): Isolation of Cell Nuclei Using Inertmacromolecules to Mimic
the Crowded Cytoplasm. Plos One. 4, 10, E7560.
Malyavantham, Kishore S. – Bhattacharya,
S. – Barbeitos, M. – Mukherjee, L. – Xu, J. – Fackelmayer, F. O. –
Berezney, R. (2008): Identifying Functional Neighborhoods within the
Cell Nucleus: Proximity Analysis of Early S-Phase Replicating
Chromatin Domains to Sites Oftranscription, RNA Polymerase II,
HP1gamma, Matrin 3 and SAF-A. Journal of Cellular Biochemistry.105, 2,
391–403.
WEBCÍM
>
Meshorer, Eran – Yellajoshula, D. –
George, E. – Scambler, P. J. – Brown, D. T. – Misteli, T. (2006):
Hyperdynamic Plasticity of Chromatin Proteins in Pluripotent Embryonic
Stemcells. Developmental Cell. 10,1, 105–16.
WEBCÍM
>
Misteli, Tom (2007): Physiological
Importance of RNA and Protein Mobility in the Cellnucleus.
Histochemistry and Cell Biology. 129, 1, 5–11.
WEBCÍM
>
Rando, Oliver J. – Chang, Howard Y.
(2009): Genome-wide Views of Chromatin Structure. Annual Review of
Biochemistry. 78, 245–271.
Székvölgyi Lóránt – Rákosy Z. – Bálint B.
L. – Kókai E. – Imre L. – Vereb G. – Bacsó Z. – Goda K. – Varga S. –
Balázs M. – Dombrádi V. – Nagy L. – Szabó G. (2007):
Ribonucleoprotein-masked Nicks at 50-Kbp Intervals in the Eukaryotic
Genomic DNA. Proceedings of the National Academy of Science of the
USA. 104, 38, 14964–14969.
WEBCÍM >
LÁBJEGYZETEK
1 Szabó Gábor akadémikus
(1927–1996) – édesapám – így fogalmazta meg ezt az örömöt: „Olvasok /
és a milliárdnyi / ember között/megtalálom/a gondolatot, / mely
összeköt. / Járhatsz / az északi / vagy déli / sarkon, / a Himalája /
ormán / vagy lent / az óceán / fenekén, / nincs lelkesítőbb, /
csodásabb kaland, / mint / rátalálni / másban / egy / benned érő / új
gondolatra…”
<
2 Az idézett irodalmak a
témakör áttekintésére közvetlenül vagy az idézett további referenciák
révén alkalmas, illetve szempontunkból gondolatébresztő közlemények,
általában nem a megfigyelések első közlései.
<
3 A magszerkezet globális,
egyszerű morfológiai vizsgálómódszerekkel is észlelhető különbségei
sokkal korábban váltak ismertté, mint az egyes gének szintjén
megmutatkozó lokális különbségek. A kromatinon zajló folyamatok
konvencionális molekuláris biológiai megközelítésének szükségszerűen
„mióp” voltára George Klein (Karolinska Intézet) már évtizedekkel
ezelőtt felhívta a figyelmet.
<
4 Az alábbi bekezdés
Werner Heisenberg A rész és az egész. Beszélgetések az atomfizikáról.
(Gondolat, 1975) 9. fejezetéből való. A szerző Niels Bohrt idézi: „Egy
élő organizmust leírhatunk először is olyan fogalmakkal, melyeket az
ember az élőlények társaságában eltöltött évezredek során alakított
ki. Ez esetben szoktunk »létről«, organikus funkciókról,
metabolizmusról, lélegzésről, gyógyulásról és hasonlókról beszélni.
Másrészt azonban megtehetjük, hogy csak az oksági folyamatok iránt
érdeklődünk. Ekkor a fizika és a kémia nyelvét fogjuk használni,
kémiai vagy elektromos folyamatokat tanulmányozunk például az
idegműködés titkait fürkészve; és feltételezzük, mégpedig szép
sikerrel, hogy a fizika és kémia törvényei – maradéktalanul érvényesek
az élő organizmusok viszonylatában is. Nos, e két szemléletmód
ellentmond egymásnak. Az első esetben ugyanis azt feltételezzük, hogy
az eseményeket a céljuk, a rendeltetésük determinálja. A másik
szemléletmód szellemében viszont azt valljuk, hogy az egyes
eseményeket az őket közvetlenül megelőző események determinálják.
Mármost merőben valószínűtlen, hogy a kétféle megközelítés csak
mintegy véletlenül vezessen nyilvánvalóan azonos eredményre.” Máshol,
ugyanebben a fejezetben, szintén Bohrt idézve: „Könnyen meglehet, hogy
a kvantummechanikában rejlő matematikai formák végtelen gazdagsága
felölelheti a biológia törvényszerűségeit is.”
<
5 Ennek a kettősségnek a
metaforájaként kívánkozik ide: „Az éjjel rászálltak a fákra, mint kis
lepkék, a levelek.” (József Attila: Eszmélet)
<
6 Az egyes
sejtféleségekben nagyságrendileg tízezernyi különböző gén fejeződik
ki, részben átfedő kombinációkban.
<
7 Az epigenetikai jelző a
génkifejeződési mintázat sejtosztódás során megfigyelhető
propagálódására utal – melynek mechanizmusairól még csak felületes
ismereteink vannak.
<
8 Talán e kérdéskör
vizsgálatát nehezíti leginkább egy „biológiai Heisenberg-reláció”:
csak nagy körültekintéssel nyerhető valódi biológiai üzenetet
tartalmazó információ a rendszerbe való jelentős beavatkozás nélkül.
<
9 Utalás Szentkuthy Miklós
hasonló című könyvére (Szépirodalmi Könyvkiadó, 1985) – a végső
összegzésre törekvés metaforájaként.
<
|