|
|
A biológia története iránt érdeklődők két jelentős
évfordulóról is megemlékeztek az elmúlt években: a svéd Carl Linné
születésének 300. évfordulójáról 2007-ben, illetve az angol Charles
Robert Darwin születésének bicentenáriumáról 2009-ben, amely
véletlenül egybeesett fő műve, a The Origin of Species első
kiadásának 150. évfordulójával. Linné méltatása elsősorban arra
összpontosult, hogy neki sikerült először rendet teremtenie az
„összevisszaságban”: az állatok és növények általa javasolt
osztályozása hosszú időn át vezérfonalként szolgált – és szolgál ma is
– a természetbúvárok számára. Mindezt
kiegészíthetjük azzal, hogy ő tette általánosan elfogadottá a kettős
nevezéktant (binominális nómenklatúra), amely két szó, a nemzetség-
(genus) és a faj- (species) név felhasználásával lát el címkével
minden élőlényt. Ez ugyan nem volt egészen eredeti ötlet,1
de ezen a téren mégis Linné munkássága számít kiindulópontnak: az
általa adott nevek a legrégebbi ma is elfogadott elnevezések a
biológiában. Darwinról talán még többet írtak és írnak manapság, mint
Linnéről, hiszen az evolúció egyik fő mechanizmusáról szóló
magyarázata, a természetes szelekcióról kifejtett elmélete döntő módon
alakította át a biológusok szemléletét és ezáltal az élet tudományát,
és igen nagy hatással volt más diszciplínák művelőire és laikusokra
egyaránt. Ő sem alkotott előzmények nélkül, de kétségen kívül
mindenkinél világosabban és elegánsabban fejtette ki véleményét, és
zseniálisan összegezte kora tudásanyagát az élővilág átalakulásáról.
Ma, a genetika és a molekuláris biológia felfedezéseinek ismeretében
válik – szinte – minden biológus számára bizonyossággá mindaz, ami
Darwin idejében még csak feltételezés volt.
Mindezt megemlékező cikkek és előadások százai
meglehetős részletességgel tárgyalták mindkét tudós esetében, de
néhány kérdés – úgy tűnik – kevés figyelmet kapott. A két évforduló
kapcsán háttérben maradt annak vizsgálata, hogy Linné és Darwin hatása
együttesen miképpen, milyen viszonylagos súllyal érvényesül a XXI.
század biológusainak gondolkodásában, szemléletük formálásában.
Összeegyeztethető-e mindaz, amivel a biológia történetének eme két
géniusza meglepte a tudományos és nem tudományos világot? Vannak-e
olyan részletek, ahol változtatnunk kell saját felfogásunkon, több
évtizedes-évszázados beidegződéseinken? Van-e olyan lényeges szempont,
amelyről kicsit megfeledkeztünk a nagy ünneplésben? Ebben a cikkben
ezekre a kérdésekre keressük a választ.
Állandóság és változás
Mondanunk sem kell, Linné és Darwin két, alapjaiban eltérő
szemléletmód képviselői – amelyek között jóval nagyobb ellentét
feszül, mint ami százévnyi eltérésből általában következik a
tudománytörténetben. Linné korában az evolúció gondolata még fel sem
merült, így ő lényegileg az élőlények változatlanságában hitt.
Zsenialitása abban mutatkozott meg leginkább, hogy szakított a kor
általános felfogásával, miszerint a természeti objektumok – ideértve a
„természetfeletti lényeket” is – egyetlen egy nagy osztályozási sorba,
a természet lépcsőjébe vagy létrájába (scala naturae) illeszkednek.
Ebben legalul vannak az élettelen dolgok (például
ásványok), amelyeket a növények csoportjai, az állatok, az ember, majd
– „természetesen” – a természetfelettiek követnek. Vagyis, mindenki
egy olyan, egymást kizáró csoportokból álló (ún. exkluzív)
hierarchiában gondolkodott, ami az egyházi méltóságok rangsorára2
emlékeztet, de jól ismert a kasztrendszer vagy a katonai rendfokozatok
példáján is. (Eme felfogás maradványa egyébként az, amikor sokan ma is
„alacsonyabb rendű” és „magasabb rendű” vagy „rangú” fajokról,
szervezetekről beszélnek.) Linné meg volt győződve arról, hogy minden
természeti objektum osztályozható, de a „természet rendjéről” más volt
a felfogása. Az osztályozás alapegységei, a fajok nemzetségekbe, a
nemzetségek rendekbe (ordo), a rendek osztályokba (classis)
sorolhatók-sorolandók, mert egy ilyen rendszer tükrözi jobban a
valóságot, és ennek segítségével sokkal hatékonyabban igazodhatunk el
a sokféleségben. (A család [familia] és sok más
kategória csak később került a rendszertani
fokozatok listájára.) Linné rendszerében az egyes csoportok
mellérendelt relációban állnak egymással: például minden nemzetség
egyenrangú, függetlenül attól, hogy milyen „fejlettségű” szervezeteket
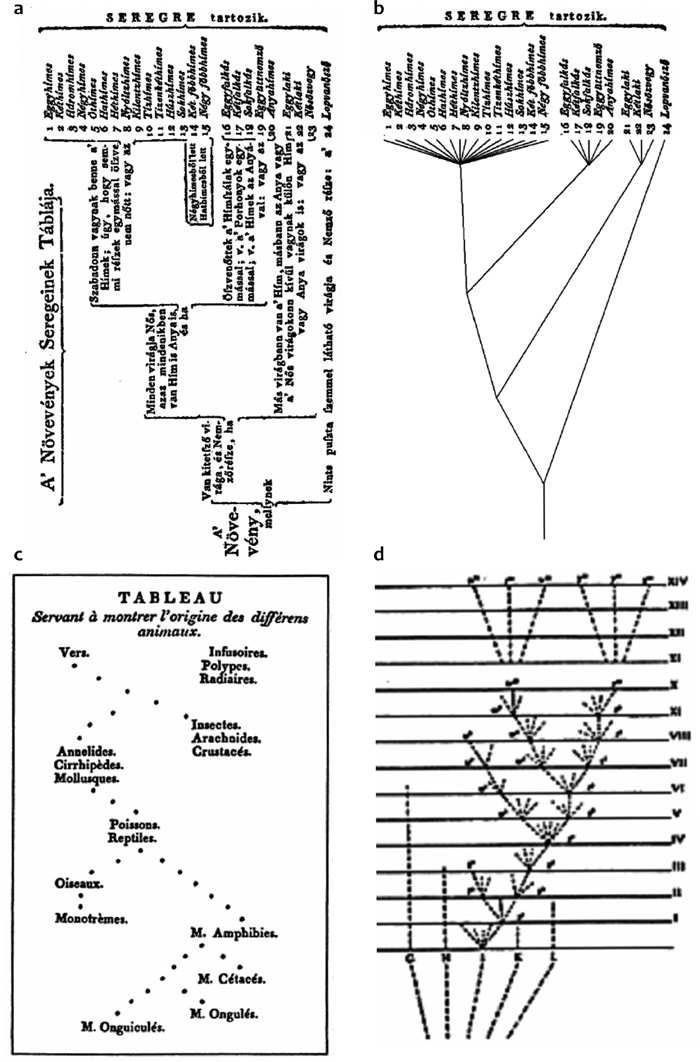
foglal magába. Matematikus szemmel nézve Linné osztályozása fa-gráfnak3
is felfogható (1/a, 1/b ábra), ami azért
érdekes, mert a fák később kiemelt fontosságot nyertek a biológiában.
Eme skatulyázós, ún. inkluzív hierarchia felállításához egyértelműen
az arisztotelészi logika adta az alapötletet (hadd utaljak csupán
olyan jól ismert fogalmakra, mint a divisio, a genus proximum és a
differentia specifica). Három különálló osztályozást alkotott Linné,
egyet a növények, egyet az állatok, és egy harmadikat pedig a kőzetek
(és kövületek!) számára. Élesen eltérő módon viszonyult az állatokhoz
és a növényekhez; míg az előbbiek csoportosítása természetes volt, és
itt-ott megegyezik mai felfogásunkkal is, az utóbbiak osztályozását
mesterségesen kiragadott bélyegekre, a porzók, illetve a termők, azaz
az ivarszervek sajátságaira alapozta. Talán kevéssé ismert, hogy ezzel
a „szexista” felfogással igencsak magára haragította kora tudományos
és nem tudományos közvéleményét egyaránt, s a reakció erejét csak a
Darwinnal szemben mutatkozó ellenségeskedéssel lehet összehasonlítani.
Botanikus kollégái felháborodottan kommentálták azt, hogy „egy nőre
akár tíz férfi is juthat” a virágban, így Linné egy ideig emigrációba
szorult, műveit pedig a Vatikán könyvtára évtizedekig nem fogadta be.
Idősebb korára Philosophia botanica című könyvében közvetve már
elismerte növényi osztályozásának túlzottan mesterséges jellegét, és
az alaktani hasonlóságokon alapuló természetes csoportosítást is
bemutatta.
Bár Linné alapvetően az állandóságra, a fajok, a
rendek és más kategóriák megváltoztathatatlanságára esküdött, azt
elismerte, hogy kereszteződéssel új fajok is létrejöhetnek. Az a
felismerés azonban, miszerint más jellegű változások a hibridizációnál
sokkalta nagyobb mértékben alakítják az élővilágot, még hátra volt. Az
evolúcióról van szó, amelynek alapgondolata – az általános
vélekedéssel ellentétben – korántsem Charles Darwin műveiben bukkant
fel először. Nagyapja, Erasmus Darwin, a francia Geoffroy
Saint-Hilaire és különösképpen Jean-Baptiste Lamarck már jóval előbb
felvetették a fajok egymásból való átalakulásának lehetőségét. Ezen a
ponton tehát akkor szolgáltatunk igazságot, ha most egy másik –
sajnálatosan mellőzött – évfordulóról is megemlékezünk: Darwin
születésének évében, azaz 1809-ben jelent meg ugyanis Lamarck
könyvében az első ábra, amely különféle állatcsoportok egymásból való
kialakulásának útvonalait vázolta fel, vagyis ez volt az első
törzsfa-jellegű ábrázolás (1/c ábra). Bár a bemutatott
kapcsolatok egyike-másika mai szemmel nézve meglehetősen naiv
feltételezéseket takar (például a tojásrakó emlősök leszármaztatása a
madarakból), szerintem Lamarck rajza egyike a biológiatörténet
legfontosabb ábráinak. Lamarck érdemtelen háttérbe szorítása
egyértelműen annak köszönhető, hogy ő még nem talált megfelelő
magyarázatot az evolúció hajtóerejére, mert ezt a szerzett
tulajdonságok utódgenerációkra történő továbbvitelében jelölte meg. A
mai szemmel nézve is igen precíz válaszra még éppen ötven esztendeig,
Darwin fő művének a megjelenéséig kellett várni.
Darwin alapvető felismerése az volt, hogy hosszú
távon, nagy időskálán nézve a változások oka a természetes
kiválogatódásban, a rátermettebbek nagyobb eséllyel történő
megmaradásában keresendő. Mindezt briliáns logikával fejti ki,
nagymértékben támaszkodva elődei és kortársai megfigyeléseire,
különösen az állatok domesztikációjával kapcsolatos tapasztalataira.
Íróasztala mellett maradva, könyveket, cikkeket és leveleket
olvasgatva azonban ő sem tudta volna megtenni a döntő lépést.
Közismert módon, a Beagle nevű vitorláson tett világkörüli utazásán
észleltek, és elsősorban a Galapagos-szigeteken tapasztaltak
erősítették meg abban a felismerésben, hogy a ma megfigyelt élővilág
pillanatfelvétel csupán és korántsem befagyott állandóság. Az a
sokféleség, amit ma látunk, a fajok populációinak összetételében
mutatkozó apró, önmagukban jelentéktelennek tűnő (ha egyáltalán
észrevehető) változások sorozatával, vagyis az evolúció révén jött
létre. A biológiát szinte sarkaiból kifordító érveinek kifejtéséhez
megdöbbentő módon mindössze egyetlen egy (!) rajzra volt szüksége
(1/d ábra) – míg manapság egy átlagos tízoldalas
tudományos cikkben van vagy négy-öt grafikon. Ennek segítségével a
fajok keletkezésének, a populációk elválásának és kipusztulásának
hipotetikus útvonalait mutatja be, ami a populációk közötti
ős–leszármazott kapcsolatok első, nyomtatásban megjelent ábrázolása.4
A mikroevolúció és makroevolúció összefüggéseit magyarázó rajzára úgy
hivatkozhatunk, mint az első evolúciós vagy filogenetikai fára, s amit
képzeletbeli galériánkban méltán helyezhetünk Linné alkotmánya és
Lamarck törzsfája mellé. Az igazság kedvéért azonban a skót–angol
származású Alfred Russel Wallace (1823–1913) nevét is meg kell
említenünk, hiszen ő is hasonló következtetésekre jutott utazásai
során, mint honfitársa. Darwin egyébként akkor döntötte el véglegesen,
hogy saját, csaknem kész kéziratos anyagával nem vár tovább, gyorsan
befejezi és kiadja A fajok eredetét, amikor Wallace hasonló tartalmú
írása hivatalos úton a kezébe került.
Egy történeti gyökerű konfliktus
Mint a cikk elején már utaltam rá, a biológusok túlnyomó többsége
egyetért abban, hogy a Földünkön megfigyelhető biológiai
sokféleség az evolúció eredménye. Minden rendelkezésünkre álló
megfigyelés, adat, számítógépes modell és tudományos érvelés emellett
szól, s maradéktalanul beilleszthető az evolúcióelmélet kereteibe, míg
az evolúció tényét cáfoló bizonyítékot még senki sem tudott
felmutatni,5 koherens elméletről nem is
szólva. Következésképpen, az élővilágot leíró és összesítő
osztályozásnak, a rendszerezés tudományának, vagyis a taxonómiának az
evolúciós folyamat elsőbbségéhez kellene alkalmazkodnia. Darwin több
helyen, így A fajok eredetében is kifejtette abbéli vélekedését, hogy
az a leginkább természetes osztályozás, amely a leszármazást veszi
figyelembe – s nem, mondjuk, a szemmel is felmérhető morfológiai
hasonlatosságot. Másik híres művében, a The Descent of Man, and
Selection in Relation to Sex (1871) fogalmazta meg ezt igazán
egyértelműen:
[The natural system,] „it is now generally
admitted, must be, as far as possible, genealogical in arrangement, –
that is, the co-descendants of the same form must be kept together in
one group, apart from the co-descendants of any other form; but if the
parent-forms are related, so will be their descendants, and the two
groups together will form a larger group”.
Vagyis: valamely ős összes leszármazottját egy
csoportba kell sorolni csakúgy, mint bármely másikét, s ha két ilyen
ős rokona egymásnak, akkor ezek összes leszármazottja együttesen
alkosson egy nagyobb taxont. Ennek az elvnek a gyakorlati
érvényesítését azonban a fentiekben vázolt történetiség, – az a tény,
hogy Linné száz évvel megelőzte Darwint – jelentősen nehezíti.
Időrendben az osztályozások jelentek meg először, törzsfák jóval
később, és az evolúció korrekt magyarázata pedig még később látott
napvilágot – míg ma már látjuk, Darwin fent idézett javaslata alapján
és minden logika szerint fordított lenne a „helyes” sorrend: a
populációk evolúciós megváltozása a háttérfolyamat, ez összegezhető
evolúciós fa formájában és az osztályozásnak az utóbbin kellene
alapulnia. Ezt az ellentétet még valahogy ki lehetne küszöbölni, de
Linnét követően, még azelőtt, hogy az evolúcióelmélet széles körben
elfogadottá vált volna, a biológusok az osztályozás minden aspektusát
szabálykönyvekben („nevezéktani kódokban”) rögzítették. A Linné-féle
hierarchia alkalmazása „kötelezővé” vált minden rendszertanos kutató
számára, s ma is „előírás”, hogy az újonnan felfedezett fajokat vagy a
hierarchia más csoportjait (ún. taxonokat) a már meglévő Linné-féle
hierarchiába kell besorolni (a kihalt formákat is). Mindez
nélkülözhetetlennek tűnik a káosz elkerülése érdekében, amint az is
alapvetően fontos, hogy minden szervezet azt a nevet viselje, amit
első leírója adott neki. Ez és a száz év „előny” viszont azt
eredményezte, hogy Linné felfogása az osztályozásról kőkeményen
bevésődött a szakmai gondolkozásba és a köztudatba egyaránt – számos,
előre nem látható következménnyel…
A múlt rekonstrukciója
Ha tehát Darwin véleményét komolyan vesszük, a linnéi hagyományokon
nyugvó osztályozásokat erőteljes revízió alá kell-kellene venni. Ezen
a ponton az olvasó joggal kérdezheti: hogyan lehetne eljárni egy
évmilliárdok óta zajló folyamat figyelembe vételével, amikor
ismereteink a jelen élővilágáról is tökéletlenek, nemhogy a régmúlt
korokéról?6 Ha a világ összes múzeumának
összes szekrénye csak fosszíliáktól roskadozna, akkor se gondolhatnánk
komolyan, hogy az evolúció útvonalait részletesen és dokumentáltan
rekonstruálni lehetne. S valóban, ajánlásait Darwin saját maga sem
tudta igazán érvényesíteni egyetlen egy rendszertani jellegű
monográfiájában, amelyet a tengerek erősen módosult ízeltlábúiról, a
kacslábú rákokról (Cirripedia) készített. Bár sok elméleti
kérdést vetett fel ennek során, lényegében véve hasonló módon
osztályozott, mint akármelyik kollégája bármely másik rendszertani
csoport vizsgálata során.
Tényleg ennyire naiv lett volna Darwin, amikor az
osztályozással szembeni kívánalmait megfogalmazta? Inkább arról van
szó, hogy ebben a vonatkozásban is előrelátó volt, mint sok mindenben,
és sejtette: javaslata előbb-utóbb – valamilyen formában –
megvalósíthatóvá válik. Kortársa és műveinek elhivatott
népszerűsítője, a jénai Ernst Haeckel (1834–1919) például igen sokat
tett azért, hogy a klasszifikáció evolúciós alapokra kerüljön. Az
igazi áttörés azonban csak a XX. század közepe táján következett be
egy német rovarász, Willi Hennig (1913–1976) munkásságának
köszönhetően. Ötlete nagyon egyszerű volt, mondhatnánk, csak éppen ki
kellett találni. Hennig úgy vélekedett, hogy az
egyes tulajdonságok nem egyformán fontosak az osztályozásban, és csak
azokat érdemes figyelembe venni, amelyek evolúciós újdonságnak
számítanak a vizsgált csoporton belül. Minél több ilyen sajátságban
egyeznek meg egyes fajok, feltehetőleg annál közelebbi rokonok. Némi
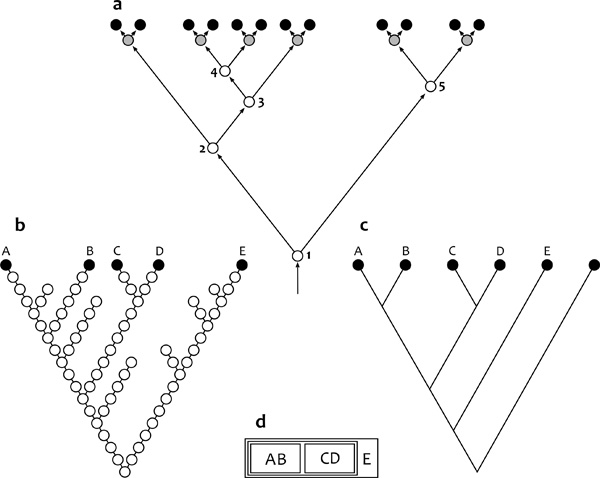
számolás után a fajok teljes rokonsági rendszerét egy fával, az ún.
kladogrammal tudjuk összesíteni (2/a ábra,
ami újabb jelölt a nagyhatású illusztrációk képzeletbeli
galériájában). Ennek végágain helyezkednek el a vizsgált szervezetek,
míg a belső szögpontok csupán hipotetikus közös ősöket képviselnek.
Hennig módszerének eredetisége tehát abban áll, hogy a ma élő
szervezetek ismeretében – vagyis anélkül, hogy kihalt rokonaikról,
őseikről bármi konkrétumot is tudnánk (!) – javaslatot tesz azok
leszármazási mintázatára.
Hennig módszere egy ideig feltűnés nélkül szunnyadt német nyelvű
publikációiban, de könyvének amerikai kiadása már komoly reakciót
váltott ki. A kladisztikai irányzat rövidesen kiszorította a 70-es
évek elején még népszerű numerikus taxonómiai iskolát, átvéve attól az
objektivitásra való törekvés igényét. A számítástechnika fejlődése
közben lehetővé tette, hogy egyre több adatot egyre hatékonyabban
lehessen a kutató döntéseitől viszonylag függetlenül és megismételhető
módon kiértékelni. Az alaktani bélyegek azonban sokszor
megtéveszthetnek bennünket: gyakori jelenség, hogy valamely
evolúciósan új tulajdonság több csoportban egymástól függetlenül is
megjelenik, megkérdőjelezve a kladogram-szerkesztés sikerességét. A
múltbéli történések sora, mint arra Emil Zuckerkandl és Linus Pauling
(1901–1994) már 1965-ben felhívták a figyelmet, sokkal jobban
megismerhető a fehérjék, illetve a nukleinsavak segítségével. A
biológusok figyelme tehát lassan az óriásmolekulák felé fordult,
hiszen építőköveik (aminosavak, illetve nukleotidok) sorrendisége
minden korábbi morfológiai adatbázisnál sokkalta nagyobb
információtartalommal bír, s ennél fogva kevésbé érzékeny a
párhuzamosságokra. A molekuláris biológia gyors
fejlődése tette lehetővé, hogy mindez ne csak elvi lehetőség legyen,
hanem a gyakorlatban is alkalmazhatóvá váljon. Ma már ott tartunk,
hogy biológus kutatók ezrei foglalkoznak kladisztikával, nemcsak
tisztán „törzsfagyártási” céllal, hanem például koevolúciós folyamatok
és biogeográfiai történések elemzésében is.7
Eredményeiket a Tree of Life nemzetközi projekt
(http://tolweb.org/tree/) egyesíti, igazolva, hogy ma már minden
eddiginél közelebb vagyunk Darwin álmának megvalósulásához, az élet
fájának rekonstrukciójához.
Az élet egyetlen fája
Az evolúcióelmélet népszerűsítésében élen járó Richard
Dawkins fejtette ki közérthető módon, hogy az evolúció egyedi és
megismételhetetlen folyamat, így annak csak egyetlen egy fa felelhet
meg hiánytalanul, s minden más ábrázolás szükségképpen tévedésekkel
terhelt. Igen fontos azonban tisztázni, hogy milyen jellegű is az a
gráf, amit egyáltalán keresni érdemes. A darwini filogenetikai fa8
populációk közötti átmenetek összegzése generációról generációra,
amelyben két pontot akkor kötünk éllel össze, ha közvetlen
leszármazási kapcsolat van a nekik megfelelő két populáció között. A
gráf tehát ős–leszármazott relációkat mutat be, és ily módon az
evolúciós folyamatok időbeli, diakrón ábrázolásának tekintendő (2/b
ábra). Hiányos múltbéli ismereteink miatt azonban biztosak
lehetünk abban, hogy ezt az igazi fát sohasem tudjuk megszerkeszteni,
ismereteink ehhez nemcsak tökéletlenek, hanem – szó szerint –
jelentéktelenek is. Emiatt, ha populációk helyett fajokban vagy
magasabb rendszertani kategóriákban gondolkozunk is, a darwini
filogenetikai fa megszerkesztése, a múltbeli történések tökéletes
rekonstrukciója elvileg lehetetlen marad. Ennek ellenére természetesen
a mégoly hiányos paleobiológiai ismeretek birtokában is megengedhetünk
olyan filogenetikai következtetéseket, mint például „a madarak
hüllőkből származtak”, vagy pedig „az egyszikűek ősei
kétszikűek voltak”, de ezeket az átalakulásokat a fenti
mikroevolúciós részletességgel sohasem tudjuk feltárni, megismerni.
Darwin törzsfájára tehát mint evolúciós modellre kell elsősorban
tekintenünk, melynek totális rekonstrukciója illúzió csupán. Más a
helyzet a kladogramokkal, mert ezek nem ős–leszármazott kapcsolatokat,
hanem az egy időben élők9 testvércsoporti
relációit fejezik ki. Két populációt vagy fajok két csoportját
testvérnek mondjuk (azaz legközelebbi rokonai egymásnak), ha őket egy
belső szögpont – ama hipotetikus közös ős – közvetlenül kapcsol össze
a gráfban. S a lényeg most következik: ha minden belső szögpont
helyesen mutatja a hozzá tartozó testvércsoporti relációt, akkor ez
azt jelenti, hogy a darwini evolúciós fa ma élőket is tartalmazó
részének a vázát (topológiáját) találtuk meg (2/c ábra). A
kladogram tehát az evolúciós mintázat sajátságos egyidejű, szinkrón
reprezentációja, s csak ebben az értelemben tekinthetjük azt
filogenetikai rekonstrukciónak: Az élet fájának megkereshető és
megbecsülhető változata voltaképpen „csak” az élet egyetlen
kladogramja lehet.
Az evolúciós fák és a kladogramok
összetévesztéséből, az általuk bemutatott relációk félreértéséből
rengeteg át nem gondolt megjegyzés és helytelen
következtetés adódik még a legkomolyabb folyóiratokban is. Nagyon
gyakori és zavarba ejtő például az, amikor az oldalsó ágra került
taxont az összes többi faj ősének tekintik (például az E-t az A-D
ősének, 2/c ábra), holott nyilvánvaló: egyik ma élő faj sem
lehet más, vele egy időben élő szervezetek bármely csoportjának
evolúciós elődje.10
A ma élő kétéltűek nem ősei a mostani hüllőknek, egyik ma ismert
kétszikű sem elődje a többieknek, a recens „majmok” – természetesen –
nem ősei az embernek és így tovább. Minden ilyen téveszmére Kevin
Omland és munkatársai találóan a primitive lineage fallacy
kifejezéssel hivatkoznak. A kladogram alapján nem lehet olyan
kérdéseket megválaszolni, mint például: melyik faj a legősibb, a
legfiatalabb, vagy éppen a „legprimitívebb”.
|
|
|
Hogyan osztályozzunk?
A legtöbb félreértés, cikkek százaiban kifejtett érvelés és a vallási
viták hevességével felérő szópárbaj azonban arra vonatkozik, hogy
miképpen osztályozzunk, figyelembe vegyük-e a leszármazást, s ha igen,
hogyan? Az osztályozás egyértelműen humán tevékenység, hiszen
objektumok egy adott halmazára sokféle osztályozás készíthető,
mindegyiket az adott célhoz igazítva (például a növényeket
felhasználásuk, földrajzi elterjedésük, életformájuk vagy éppen a
nevük kezdőbetűje szerint is osztályozhatjuk). De milyen legyen akkor
az élet „valódi”, rendszertankönyvekbe való egyetlen, kitüntetett és
objektív osztályozása? Miután az evolúciós folyamat egyedi, a kutató
tudatától és mindennemű befolyásoltságtól leginkább mentes osztályozás
az lesz, ami e folyamat összegzésén, a törzsfán alapszik. S valóban,
egy törpe kisebbségtől eltekintve a biológusok zöme ma már egyetért
abban, hogy a rendszertani csoportok elkülönítésében – Darwin
nyomdokain – a közös leszármazás legyen a fő szempont. Az igazi
nézeteltérések viszont ezután kezdődnek…
A Linné-féle osztályozás statikus és igen merev
konstrukció: minden egyed egy fajhoz, minden faj egy adott génuszhoz
tartozik és így tovább. Ezen csoportok között definíciószerűen éles a
határ – átfedés legfeljebb a hibridfajok esetében jelentkezik, más
átmenetet a taxonómia nem ismer. Linné inkluzív hierarchiájában
„felfelé haladva” a morfológiai szakadék szélesedik, két génusz
elkülönítéséhez több eltérés kell, mint két faj elválasztásához, a
rendek elhatárolása nagyobb taxonómiai
különbözőséget tételez fel, mint a családoké, és így tovább. Ezzel
szemben az evolúció maga a folytonosság, és ha egyes populációk
hosszabb ideig változatlannak tűnhetnek is, taxonómiailag értékelhető
váltás általában nem következhetett be hirtelen egyik generációról a
másikra.11 Különösen a fajnál magasabb
rendszertani kategóriák esetében tekinthetjük ezt kizártnak. Márpedig
ha a Linné-féle hierarchiát akarjuk alkalmazni az összes valaha élt és
ma élő szervezet együttes osztályozására – a mai taxonómusok ui. ezt
teszik, Linnével ellentétben! – akkor egy paradox helyzet áll elő:
egyes múltbéli populációk között húzható faji határnak egybe kell
esnie a génusz-, család-, rend-, osztály- és törzsi szintű
elhatárolással! Nyilvánvaló azonban, hogy például az a
tulajdonságcsoport, ami a ma élő zárvatermőket a nyitvatermőktől
élesen elválasztja (például a kettős megtermékenyítés, a magház, a
vízszállító csövek) nem két generáció között alakult ki, amint azt a
szigorú hierarchikus osztályozás feltételezi, hanem hosszas evolúciós
folyamat eredménye (Tod Stuessy szerint a nyitvatermők és zárvatermők
közötti evolúciós átmenet akár százmillió évig is eltarthatott).
Rengeteg egyéb esetet is felsorolhatnánk (a biológia bővelkedik
ezekben), így csak a halak és a négylábúak, vagy a hüllők és a madarak
példáját emeljük még ki. A Linné-féle hierarchia alkalmazása kihalt
fajokra, a mában tapasztalt hierarchikus mintázat visszavetítése a
múltba, azaz egy diakrón típusú linnéi osztályozás tehát minimum
ellentmondásos az evolúciós folytonosság, a változás miatt. Mindezt
közvetve Darwin is felismerte, amikor megjegyezte,
hogy a taxonok elkülönítése – az őslénytannal foglalkozók „nagy
szerencséjére” – a fosszilis anyagban mutatkozó hiátusok miatt
lehetséges csupán – márpedig a fosszilis anyag egy olyan sajthoz
hasonlítható, amely szinte csak lyukakból tevődik össze. Tudomásom
szerint Roy Albert Crowson (1914–1999) brit rovarász12
vetette fel először azt, hogy a fenti okokból a linnéi osztályozás
csak az egy időben élő szervezeteket összesítse. De ne feledkezzünk
meg a kihaltakról sem, és készítsünk külön-külön klasszifikációkat
legalább a földtörténeti időszakok szerint, ha finomabb léptékben ez
már nem is megy. Vagyis, az osztályozás célszerűen csak szinkrón
lehet. Crowson javaslata azonban nem talált követőre, holott teljesen
összhangban van Linné elképzeléseivel, hiszen a nagy svéd tudós is –
lényegében véve – az egy idősíkban élőkre alkotta meg hierarchikus
rendszerét. Mindebből logikailag következik egyes rendszertani iskolák
(például az evolutionary taxonomy) fő problémája: elismerik ugyan az
evolúció szerepét a földi biodiverzitás kialakításában, de furcsa
módon az evolúciós folytonosságot nem veszik igazán komolyan, és
ráadásul nem kötődnek a valódi linnéi hagyományhoz, azaz az
egyidejűséghez és állandósághoz sem.
A kiutat, és ezt az olvasó a fentiek alapján már ki
is találhatja, a kladisztika kínálja. A linnéi hierarchikus
osztályozás nyilván nem vagy csak nehezen és rengeteg bonyodalommal
váltható fel mással, így ezt a legtöbb biológus meg szeretné tartani.
Ez természetesen érthető és célszerű is, de Darwin ajánlását is jó
lenne figyelembe venni, hiszen ez ad elméleti hátteret az
osztályozáshoz. Mindehhez a Hennig javasolta kladisztikai módszerek és
azok molekuláris adatok feldolgozásához illesztett szofisztikált
változatai jelentik a módszertani bázist. A kladogramok pedig
viszonylag könnyen átalakíthatók egy Linné-féle hierarchiába, oly
módon, hogy minden taxonban közös leszármazásúakat (és csak azokat)
egyesítünk (lásd a 2/d ábra egyszerű példáját). A kapott
klasszifikáció így megfelel Crowson ajánlásainak is, vagyis: az
élővilág linnéi osztályozása a darwini kritériumok alapján teljesen
reális célkitűzés, ha figyelmünket a ma élő szervezetekre
összpontosítjuk, s a kladisztika eredményeiből indulunk ki.
Ma már példák serege igazolja, hogy a többség –
kimondva, kimondatlanul – a fenti utat követi. Figyelemre méltó, hogy
számos esetben a tradicionális taxonómia csoportosítása a molekuláris
elemzések tükrében is megállja a helyét. Törzsek, osztályok, rendek és
családok sokaságáról bizonyosodik be, hogy a makromolekulák sem
mondanak mást, mint a morfológiai tulajdonságok. A növények köréből a
zárvatermőket, az egyszikűeket és a fészkeseket említhetjük, mint
ahogy a puhatestűek, a gerincesek, az emlősök, a madarak és a lepkék
is jól körülhatárolható csoportokat jelentenek az élet egyetlen igazi
kladogramján. Sok esetben azonban már a korábbi morfológiai és a
későbbi molekuláris kladisztikai eredmények is megváltoztatják korábbi
elképzeléseinket. Kiderül például, hogy az állatok számos törzse,
amelyekről azelőtt senki sem gondolta volna, hogy filogenetikai
rokonság fűzné össze őket (fonálférgek, kerekesférgek, ízeltlábúak)
egy nagy ágat jelentenek az állatvilág törzsfáján (Vedlők –
Ecdysozoa), mert mindannyian vedléssel szabadulnak meg kinőtt
kültakarójuktól. Azt se sejtette senki, hogy a kétszikűek egy jól
körülhatárolt csoportja (Eurosid I) egyesíti magában mindazokat
a növénycsaládokat, amelyekben nitrogénkötő baktériumokkal
szimbiózisban élő fajokat találunk (például a pillangósok,
nyírfafélék). Ezek a korábbi rendszerekben egymástól igen távol
voltak. A kladisztika módszerei mutatnak tehát rá olyan
összefüggésekre, amelyek egyébként homályban maradtak volna. És amikor
a morfológia totális csődöt mond, mert nincs olyan közös bélyeg,
melynek alapján egyáltalán lehetséges volna az összehasonlítás, csak a
molekulák segíthetnek. Így derül ki sok élősködőről, hogy hova is
tartozik valójában. Azelőtt nem tudtuk, hogy az egysejtű
nyálkaspórások (Myxozoa) fajai voltaképpen a csalánozók
rokonsági körébe sorolhatók, azaz egysejtű létük másodlagosan alakult
ki parazita életmódjuk eredményeként. Persze vannak olyan esetek is,
amikor a közvélekedés vonakodva fogadja el az új eredményeket. Például
„nehezen emészthető” a klasszikus rendszertanon nevelkedtek számára
az, hogy a hüllők már nem taxon, mert közös ősük egyben a madarak
közös őse is volt. Ezen nem az segít, hogy a madarakat is (egyes
javaslatok szerint) hüllőnek fogjuk fel, hanem az, hogy a hüllők ma
élő nagy csoportjait (a gyíkokat, kígyókat, teknősöket és
krokodilokat) külön-külön olyan rangra emeljük, mint a madarakat
(osztály a gerinceseken belül). A „hüllő” kifejezést pedig megtartjuk
egy olyan fejlődési szint számára, amelynek legfeljebb egy éles
határok nélküli diakrón osztályozásban lenne értelme. Ugyanígy jár a
halak vagy akár a kétszikű növények csoportja is, nem is beszélve a
mohákról és a harasztokról.
Hogyan nevezzük el az élőlényeket?
Végezetül egy olyan témát is érdemes megvizsgálnunk, amelyben első
látásra úgy tűnik, Linnének és Darwinnak nem sok köze van egymáshoz:
az élőlények elnevezésének kérdéseit. Az egységes nómenklatúra
rendkívül fontos a biológiában, hiszen enélkül nem tudnánk, hogy miről
beszélünk, mit vizsgálunk. Annak, aki egész kutatói pályáján csak az
Arabidopsis thaliana nevű kis keresztes virágú növénnyel foglalkozik
(ez ui. kedvelt alanya a molekuláris genetikai vizsgálatoknak, és ma
már igen sokat tudunk róla) éppen olyan fontos a nevezéktan, mint a
fajok százaival sőt ezreivel foglalkozó trópusi erdő-ökológusnak.
Ebben a tekintetben nincs közöttük különbség, az eltérő fajszám
ellenére sem. Ma már tisztán látjuk azonban, hogy a kettős név
bevezetése – ellentétben az inkluzív hierarchiával – óriási tévedés
volt, bár ezt sokan még mindig Linné egyik fő érdemének fogják fel. A
bajok forrása ott keresendő, hogy Linné az osztályozás alapegységeként
a fajokat jelölte meg, mint ahogy ma is tesszük, viszont a
nómenklatúra alapegysége a génusz! A fajnevek (például vernalis,
communis, aquatica vagy akár thaliana) önmagukban semmit sem érnek, ha
nem tesszük eléjük a nemzetség nevét. A génusz viszont már egy faj
feletti taxon, és az, hogy egy faj milyen nevet viseljen, ily módon
nem független a klasszifikációtól. Persze Linné döntése – a saját
szempontjai szerint – akár érthető is, mert meg volt győződve arról,
hogy azt az osztályozást „találta meg”, ami eleve teremtetett – vagyis
nemcsak a fajok állandóak, hanem majd az ő klasszifikációja is örök
érvényű marad. Az utódok viszont korántsem gondolkoztak így, hiszen
már az evolúcióelmélet megjelenése előtt alaposan átrendezték Linné
rendszerét, a kettős neveket is – és az „átdolgozás” azóta is
szüntelenül tart. Annak a megítélése ugyanis, hogy mely fajok
tartozzanak egy nemzetségbe már jóval nehezebb probléma, mint a faji
identitás keresése. Nem minden génusz olyan egyértelmű (azaz
„természetes”), mint mondjuk a tölgyeké (Quercus) vagy a
rózsáké (Rosa), melyeknél viszont inkább a fajok elkülönítése
okozza a fő problémát. A kettős nevezéktan alkalmazása azt
eredményezte, hogy vannak olyan fajok, amelyek már három-négy, sőt
több génuszt is megjártak a taxonómusok átsoroló tevékenységének
eredményeként, így az eredeti név adója sem ismerne rá. Ha Linnének
megmutathatnánk az Eleocharis palustris, Epipactis palustris és
Rorippa palustris neveket, és közölnénk vele, hogy ezeket a
fajokat ő írta le először, akkor bizony fogalma se volna arról, hogy
melyik palustris mit is rejt valójában (ui. eredetileg ezeket a
Scirpus, a Serapias, illetve a Sysimbrium génuszokba
sorolta be 1753-ban). A rendszertan tehát meglehetősen kaotikus
önmagában, az evolúciós szemlélet nélkül is. Ha minden fajt egyedi név
jelölne, s nemcsak a faj feletti taxonokat, akkor ez a probléma fel
sem merülne. Ugyanakkor a kettős nevek már annyira beépültek minden
biológus tudatába, könyvek és cikkek százezreibe, hogy bármiféle
lényeges változtatás lehetetlen lenne. Azon mindenesetre érdemes
elgondolkodni, hogy a génusz szintet esetleg törölhetjük a
rendszertani rangok sorából, s a kettős fajneveket a továbbiakban már
érintetlenül hagyjuk az osztályozás során.
Az utókor Linné hibáját egy még rosszabb döntéssel
tetézte, ami történetileg persze szintén érthető. Hosszú ideig ugyanis
az élővilágot két fő részre bontották, növényekre
és állatokra, melyeket két külön diszciplína, a botanika és a zoológia
vizsgált (ennek persze máig ható hatása van például az egyetemi
tanszékek megnevezésében). A két tudományág idővel jelentősen
elszakadt egymástól, és külön – egymástól független – nómenklatúrai
szabályrendszert állított fel magának.13
Az élőlények elnevezése tehát nem egységes, amelynek időnként igen
furcsa következményei vannak. Több száz génusz név egyformán jelöl – a
szabályok szerint érvényesen – állatot és növényt egyaránt (írjuk be a
Google képkeresőjébe a Prunella vagy Arenaria szavakat, s hasonlítsuk
össze, mik is tartoznak „ugyanabba” a génuszba). A zűrzavart csak
kismértékben – de elviekben annál súlyosabban – fokozza, hogy
tucatnyinál több esetet ismerünk, amikor a binomen mindkét része
azonos egy növény és egy állat esetében. Ezt felismerve a
bakteriológusok megalkották saját nómenklatúrájukat
a prokarióta élőlények számára – de ezzel nem csökkentették, hanem
csak növelték a káoszt. Evolúcióbiológiai értelemben ugyanis távolról
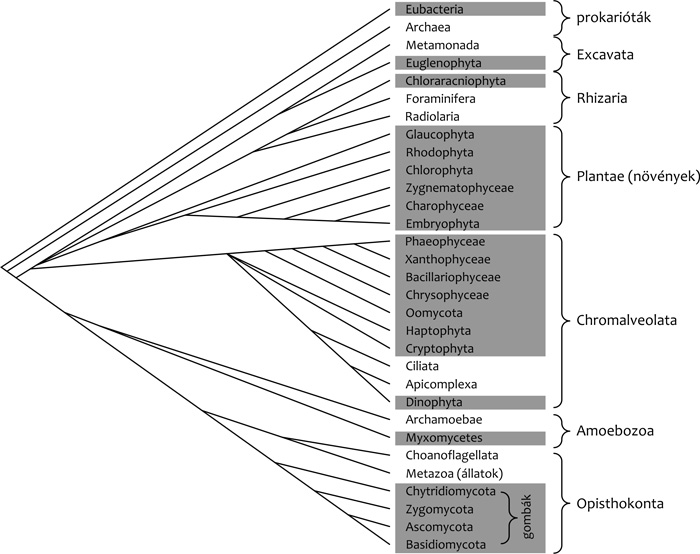
sem felel meg a valóságnak a fenti hármas felosztás, s ma már látjuk,
a növénytani nómenklatúra olyan szervezetek esetében tart igényt a
névadásra, amelyek teljesen különböző származásúak
(3. ábra). Legfeltűnőbb a gombák esete,
melyeket ma is a botanikai szabályok szerint kell elnevezni, holott az
állatokhoz sokkal közelebb állnak, mint a növényekhez. Bár voltak
próbálkozások a nevezéktan egységesítésére (például a BioCode), de
mindegyik eredménytelen maradt. Míg a tudomány más területein a „kőbe
vésés” szinte teljesen kizárt, a taxonómusok mindmáig keményen
ellenállnak minden alapvető változásnak a régi nevek „stabilitása” és
szabályaik állandósága érdekében, nem ismerve fel azt, hogy éppen
ezzel okozzák a legtöbb gondot minden evolúciós szemléletű
biológusnak.
Epilógus
Linné és Darwin, Hennig és Crowson – és sok ezer biológus kutató –
munkásságának közös tanulsága csak egy lehet: az evolúció tudománya és
az osztályozás tudománya (a rendszertan) nem választhatók el
egymástól! Egyik sem érthető meg igazán a másik nélkül, s ezt csak oly
módon tudatosíthatjuk a jelen, s főleg a jövő biológus generációi
számára, ha oktatásukat középiskolai és egyetemi szinten is
integráljuk. Így alakulhat csak ki az a fajta biológustársadalom,
amely nem részterületek elefántcsont-tornyából tekint a többiekre, s
nem értetlenkedve-gyanakodva figyeli a biológiai tudomány fejlődését –
ha egyáltalán figyeli – hanem képes az új eredmények szintetizálására,
önnön szemléletének folytonos megújítására is. Mindezt annak érdekében
tegyük, hogy valóra váltsuk minden biológus közös álmát, az élet
fájának minél pontosabb megszerkesztését, s ezáltal a rendszerezés
evolúciós alapjainak tisztázását.
Kulcsszavak: evolúció, Élet Fája, hierarchia, kladisztika,
nómenklatúra, osztályozás, taxonómia
IRODALOM
Avise, J. C. – Nelson, W. S. – Sugita, H.
(1994): A Speciational History of “Living Fossils”: Molecular
Evolutionary Patterns in Horseshoe Crabs. Evolution. 48, 1986–2001.
Blunt, W. (2002): Linnaeus. The Compleat
Naturalist. Princeton University Press, New York
Cracraft, J. – Donoghue, M. (eds.) (2004):
Assembling the Tree of Life. Oxford University Press, Oxford
Crowson, R. A. (1970): Classification and
Biology. Heinemann Educational, London
Darwin, Charles (1859): On the Origin of
Species. Murray, London
Darwin, Charles (1871): The Descent of
Man, and Selection in Relation to Sex. Vol. 1. Murray, London
Darwin, Charles (1955): A fajok eredete.
(fordította Mikes Lajos) Művelt Nép, Budapest
Dawkins, Richard (1994): A vak órásmester.
Akadémiai–Mezőgazda, Budapest
Dawkins, Richard (2007): Az Ős meséje:
Zarándoklat az élet hajnalához. Partvonal, Budapest
Dawkins, Richard (2009): A legnagyobb
mutatvány: Az evolúció bizonyítékai. Nyitott Könyvműhely, Bp.
Dayrat, B. (2005): Ancestor–Descendant
Relationships and the Reconstruction of the Tree of Life.
Paleobiology. 31, 347–353.
Ereshefsky, M. (2001): The Poverty of
Linnaean Hierarchy: A Philosophical Study of Biological Taxonomy.
Cambridge University Press, Cambridge
Hennig, W. (1966): Phylogenetic
Systematics. University of Illinois Press, Urbana
Lamarck, Jean-Baptiste (1809): Philosophie
zoologique, ou Exposition des considérations relatives à l’histoire
naturelle des animaux. Paris
WEBCÍM >
WEBCÍM >
Omland, K. E. – Cook, L. G. – Crisp, M. D.
(2008): Tree Thinking for All Biology: The Problem with Reading
Phylogenies As Ladders of Progress. BioEssays. 30, 854-867.
Padian, Kevin (1999): Charles Darwin’s
Views of Classification in Theory and Practice. Systematic Biology.
48, 2, 352–364.
WEBCÍM >
Podani János (2009): Taxonomy Versus
Evolution. Taxon. 58, 1049–1053.
Podani János (2010): Taxonomy in
Evolutionary Perspective. Synbiologia Hungarica. 6, 1–42.
WEBCÍM >
Tuba Zoltán – Szerdahelyi T. – Engloner A.
– Nagy J. (szerk.): Botanika II. Rendszertan. Nemzeti Tankönyvkiadó,
Budapest, 219–236.
Stuessy, T. F. (2004): A
Transitional-Combinational Theory for the Origin of Angiosperms.
Taxon. 53, 3–16
Zuckerkandl, Emil – Pauling, Linus (1965):
Molecules As Documents of Evolutionary History. Journal of Theoretical
Biology. 8, 357–366.
LÁBEJGYZETEK
1 A svájci Gaspard Bauhin
(1560–1624) javasolta először a kettős elnevezéseket a rendszertanban.
<
2 A hierarchia görög
eredetű szó, jelentése kb. „szent hatalom”.
<
3 A fa olyan szögpontokból
és élekből álló matematikai konstrukció, vagyis egy olyan speciális
gráf, amelyben bármely szögpontból el lehet jutni bármelyik másikba,
de csak egyetlenegyféleképpen.
<
4 Darwin jegyzetfüzeteiben
természetesen már korábban is szerepeltek hasonló rajzok, a legelső
valószínűleg 1837-ből.
<
5 Híres anekdota J[ohn]
B[urdon] S[anderson] Haldane-ról, a XX. század egyik legjelentősebb
biológusáról, aki arra a kérdésre, hogy mikor inogna meg benne az
evolúciós meggyőződés, a következőképpen válaszolt: „Fossil rabbits in
the Precambrian” vagyis akkor, ha egy nyúl-fosszília kerülne elő a
prekambrium rétegeiből. A földtörténetben kevésbé jártas olvasó
számára jegyezzük meg, ez valóban igen érdekes lelet lenne, hiszen a
prekambrium, melyben kizárólag tengeri lények éltek, 540 millió évvel
ezelőtt véget ért, míg az emlősök a triászban jelentek meg kb. 225
millió évvel ezelőtt, a szárazföldön. Ezzel a rövid idézettel
szeretném jelezni kissé ironikusan, hogy a cikk további részében az
evolúció lényegével kapcsolatos kétkedésekkel nem kívánok foglalkozni.
<
6 Az eddig leírt fajok
száma valamivel több mint másfél millió, ugyanakkor a
legkonzervatívabb becslések szerint is még legalább ennyi ismeretlen
faj vár felfedezésre (mások szerint sokkal több, akár száz millió
is!). A fosszilisan ismert fajok száma csak kb. negyedmillió.
<
7 A kladisztika
alkalmazható a biológián kívül is, ha a történetiség lényeges, mint
például a nyelvek és középkori szövegek evolúciójának elemzésében.
<
8 Darwin már-már költői
színvonalú összegzése az élet fájáról A fajok eredete 6. kiadásában:
„The affinities of all the beings of the same class have sometimes
been represented by a great tree... As buds give rise by growth to
fresh buds, and these if vigorous, branch out and overtop on all sides
many a feebler branch, so by generation I believe it has been with the
great Tree of Life, which fills with its dead and broken branches the
crust of the earth, and covers the surface with its ever branching and
beautiful ramifications.” „Egy osztály valamennyi élőlényének
rokonsági kapcsolatait néha nagy fa formájában szokták
ábrázolni...Amint a rügyek növekedésükkel friss rügyeket hoznak létre,
amelyek ha életerősek, minden irányban szétágaznak, és a gyöngébb
ágakat túlnövik, ugyanez a helyzet, véleményem szerint az élet nagy
fájával is, amely a föld kérgét halott és letört ágaival tölti meg, a
föld felszínét pedig örökké tovább ágazó gyönyörűséges elágazásaival
borítja el.” (Darwin, 1955)
<
9 Egyes speciális
esetekben fosszilis és recens fajok együtt szerepelnek a kladisztikai
elemzésben. Ám vigyázni kell, mert ekkor egy potenciális ős
testvércsoportként szerepelhet, s emiatt nehéz a kladogram
értelmezése.
<
10 Az evolúciós
folyamatok nem „állnak le” még akkor sem, ha ezt morfológiai
változások nem kísérik. Idevág az „élő kövületek” problémája, amelyek
látszólag hosszú ideje változatlanok, de valójában lényeges
molekuláris megváltozások állnak a háttérben, mint például a
tőrfarkúak (Limulus) esetében ez már közvetve be is
bizonyosodott.
<
11 Egyes biológusok
korábban felvetették a hirtelen nagymérvű változások, a szaltációk
lehetőségét is, de megmutatható, hogy ez már csak populációgenetikai
okok miatt sem képzelhető el. A legnagyobb ugrás, ami egyik
generációról a másikra lehetséges, az kb. faji szintű, s létrejöhet
hirtelen bekövetkező kromoszómaszám-változás hatására vagy
hibridizációval.
<
12 „Bogaras” ember volt,
míg Hennig a kétszárnyúak körül „legyeskedett”.
<
13 Csak egy példa: a
botanikusokkal ellentétben a zoológusok sok nevet kétszer mondanak,
kétszer mondanak (például: Vulpes vulpes – róka, Pica pica – szarka),
ami jól mutatja a kettős elnevezések időnkénti redundáns jellegét is.
<
|
|