|
|
Pléh Csaba összefoglalója megmutatja, hogyan lehet
kevés adatból, nagyon sok ésszel később is érvényes megállapításokat
tenni a nyelv biológiai alapjairól. Az én helyzetem
könnyebb, arra világítok rá, hogy rengeteg biológiai adat alapján,
nagyon kevés ésszel is milyen sokat tudtunk meg a madarak
énektanulásáról. Dolgozatomban egy rövid áttekintés után elsősorban a
legfrissebb eredményeket kívánom bemutatni. A régebbi eredmények
ismertetésekor összefoglaló munkákra hivatkozom, eredeti közleményeket
az új fejlemények leírásánál citálok.1
Fentről le modellek
Az idegtudományi kutatásokat messze megelőzve, az énektanulás terepi
és kísérletes vizsgálatából kirajzolódott már az ének elsajátításának
elvi modellje, ami később kicsit, egyesek szerint nagyon módosult.
Elsősorban Peter Marler megfigyeléseiből tudjuk, hogy az énekesmadár
fajok énekében kimutathatók a fajra jellemző (fonológiai és/vagy
szintaktikai) univerzálisok, ugyanakkor a különböző populációk éneke
különbözhet, és ezek a felismerhető dialektusok egyedszintű tanulással
adódnak át. Kontrollált laboratóriumi vizsgálatok bizonyították, hogy
a teljes izolációban nevelt madár is énekel, a dallam néha torzítva,
de tartalmazza a fajspecifikus univerzálisokat, ugyanakkor az izolált
énekből hiányoznak a különleges díszítések, az egyedi jellemzők. Ha
két dallamot hallhat a tanulási fázisban, akkor a fiatal madár a saját
fajára jellemzőt választja ki, és tanulja meg. Mindebből Peter Marler
arra következtetett, hogy a madár rendelkezik egy nemtanult
fajspecifikus mintázattal, templáttal.
Ha a fiatal madár nem hallhatja saját énekét a
gyakorlási fázisban, akkor a fajspecifikus mintázat egyáltalán nem
vagy nagyon rontott formában jelenik meg. A természetes énektanulás
ezek szerint azzal kezdődik, hogy a madár összehasonlítja a hallott
dallamokat a veleszületett (igazából „velekelt”-et kéne írni)
templáttal, a kiválasztott dallamot megtanulja, és a gyakorlás során a
saját éneket a tanult templáttal hasonlítja össze. A gyakorlás a
csecsemők gügyögésére funkcionálisan emlékeztető elődallamokkal
kezdődik, ezek plasztikusan változnak, míg a saját ének hasonlóvá nem
válik a tanult dallamhoz, illetve míg az idegrendszeri érés be nem
zárja a tanulás időablakát, a szenzitív periódust. Amikor az ének
eléri végleges formáját – kikristályosodik – a madárnak többé nem kell
saját énekét hallania ahhoz, hogy változatlan formában énekeljen. A
tanult templát rögzítése a szenzoros fázis, a gyakorlás közbeni
tökéletesítés a szenzomotoros fázis. Úgy tűnt tehát, hogy bár a
részletekben különbözhetnek az egyes fajok, az énektanulás
folyamatának lényege egy univerzális templátmodellben leírható.
Apró részletek
A korai etológia hadat viselt a behaviorizmus ellen, sok érvük között
a két legfontosabb módszertani kritika az volt, hogy a behavioristák
semmit sem tudnak az állatokról, mert ötven éven át mindössze öt fajt
vizsgáltak, és semmit sem tudnak a természetes viselkedésről, mert
állataikat mesterséges környezetben tanulmányozzák. Az éneket kutató
etológusok azonban ugyanilyen bűnbe estek. Több mint négyezer
énekesmadár fajt ismerünk, ezek közül gyakorlati szempontok miatt
mindössze öt-tíz fajt vizsgáltak, legintenzívebben egyetlenegyet, a
zebrapintyet, és ezt a fajt sem természetes körülmények között, hanem
laboratóriumban.
Laboratóriumban a dallamokat magnetofonról halló
fiatal madár valóban a saját fajra jellemző éneket választja, azonban
ha az idegen fajú tutor élő madár, akivel a fiatal szociális
kapcsolatban áll, akkor az idegen faj énekét tanulja meg. Mondhatni, a
madarak is nehezebben győzik le a kognitív disszonanciát
távoktatásban, mint személyes példák alapján. A szociális tanulás nem
csak az idegen dallam elfogadásában plasztikus, hanem abban is, hogy
később zárul le a szenzitív periódus.
A könnyen tartható és ezért sokat vizsgált
zebrapinty az összehasonlító kutatások alapján nem tekinthető átlagos
énekesnek. A zebrapintynél a tutor dallamának megtanulása (szenzoros
fázis) és a gyakorlás (szenzorimotoros fázis) átfedő, sok fajnál
azonban teljesen elkülönül (például háziveréb). A zebrapinty életében
csak egyszer tanul, sok faj azonban minden évben új dallamokat sajátít
el (például: kanári, örvös légykapó). A zebrapinty egyetlen és
egyszerű dalt tanul, sok faj több és komplex dallamot sajátít el, akár
olyan hangmintázatot is, ami a természetben nem fordul elő (például a
lantfarkú madár utánozza az autóriasztó, a láncfűrész vagy a
fényképezőgép hangját).
A természetes körülmények között végzett
vizsgálatok során az is kiderült, hogy sok fajnál a sokféle megtanult
ének közül a hímek idősebb korban szelektálnak, bizonyos dallamokat
bár megtanultak, később nem adnak elő. Ez a jelenség összefügghet
azzal, hogy az ének kiválasztása a kontextustól függ. Ha a szomszéd a
territórium határán énekel, a hím fokozhatja a konfliktust, ha
ugyanazzal a dallammal vagy a szomszéd számára ismeretlen énekkel
válaszol, de kifejezheti békés szándékát, ha a szomszéd repertoárjából
választ dallamot, mintegy jelezve, hogy ismeri a kedves szomszédot. A
madarak énektanulása és az ének használata tehát jóval plasztikusabb,
mint azt régebben feltételezték, de tegyük hozzá, még a viselkedés
szintjén sem teljes a kép, mert a vizsgálatok zömét a fejlett országok
kevésbé fejlett madarain végzik. A trópusi fajok között sok van,
amelyekben a tojók is énekelnek, vannak, amelyek egész éven át
tanulnak, és számos más jellemzőben különbözhetnek a mi madarainktól.
A hangszer
Sejthető volt, hogy az énektanulás idegi mediációja nem lehet
egyszerű, mert a hallott mintázat szenzoros reprezentációja nem
topografikus, és az éneklés motoros parancsmintázata sem az.
Feltételezhető, hogy a szenzoros és motoros kód nagyon absztrakt és
egymással nagyon szoros kapcsolatban, talán átfedésben dolgozik, amit
emlősöknél csak nemrég sikerült kimutatni (Mátyás et al., 2010)
A kutatók szerencséjére maga a hangképző szerv
viszonylag egyszerű. Az énekesmadarak a szirinxben, tehát a két
főhörgő találkozásánál egy kétoldali páros szervvel adnak hangot. A
hangot a szirinxbe betüremkedő két-két nyelvecske rezgése kelti. A
hang alaptulajdonságait, amint ezt számítógépes modellel igazolni
lehetett, lényegében két tényező határozza meg: a nyelvecskék
feszessége és a respirációs nyomás. A mozgató izmok ún. szupergyors
rostok, melyek 250 Hz frekvenciás összehúzódási ritmusa nagyon gyors
váltásokat tesz lehetővé. Az énekesmadarak légcsövének szerkezete
eltérő lehet az ivarok között, és ezt a célszerszámot a hímek képesek
a hang alapfrekvenciájára hangolni (Riede et al., 2010). További
modulációt tesz lehetővé a nyitott csőr szögének változtatása. Nem
várt, de utólag kézenfekvő megoldás, hogy a madarak a két hangképző
szervüket egymástól függetlenül is tudják működtetni, aminek
köszönhetően duettet énekelhetnek saját magukkal, és nagyrészt ez a
magyarázat a madárdal nem lineáris trükkjeire
(frekvenciaugrás, szubharmonikusok, bifonáció, determinisztikus káosz)
is.
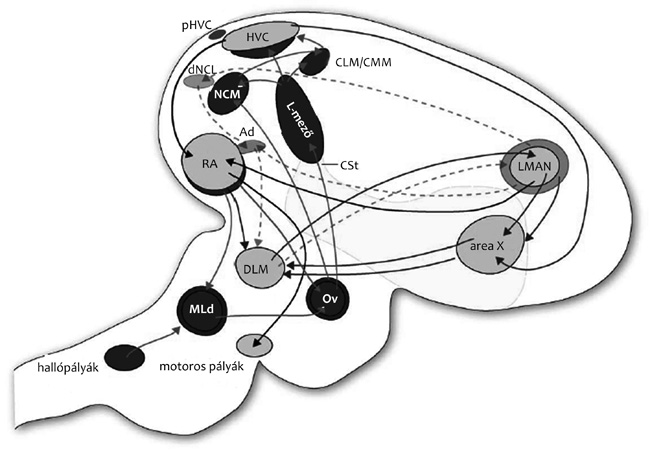
Éneklő pályák az agyban
A feladat most már csak annyi, hogy megértsük, miként lesz a hallott
énekből memórianyom és ebből éneket produkáló motoros parancsmintázat!
A fülből érkező információ alacsony szintű processzálást követően a
talamikus ovoidalis magon (Ov)2 keresztül
jut a nagyagyi hallóterületekre. Az utóbbi években
több, független bizonyíték alapján tudjuk, hogy a korábbi
elképzelésekkel szemben az énekhang feldolgozásában az elsődleges
hallóterületen (L-mezők) kívül az NCM- és CMM-magnak is fontos szerepe
van (áttekinti Bolhuis et al. 2010). Kapcsolatrendszerük és funkciójuk
alapján ezt a két magot sokan a Wernicke-terület analógjának tekintik.3
Az ének egyes hangjai tonotopikus mintázatot generálnak az NCM-ben. A
mintázat egyik különlegessége az, hogy az NCM mesterséges monotonikus
hangra nem reagál, másrészt a komplex énekhang által
generált mintázat nem vezethető vissza a részek mintázatára.
Zebrapintyek felnőtt korukban is preferálják a tutor (természetes
körülmények között az apa) énekét, a preferencia azonban megszűnik az
NCM ledálásával. Ez és más adatok arra utalnak, hogy az NCM lehet az
elsődleges tárhelye a megtanulandó éneknek. A hallott ének következő
állomása a HVC4-mag. A HVC léziója néma
éneklést okoz, a hím megfelelő helyzetben, megfelelő pózban, megfelelő
mozgásmintázattal nem hallat hangot. Az énektanulást megelőzően a HVC
térfogata jelentősen megnövekszik, mert a területre új neuronok
vándorolnak. Fernando Nottebohm ezen felfedezése nagyon jelentős volt,
mert elsőként bizonyította, hogy funkcionális kapcsolatot teremtő új
neuronok születnek felnőtt gerinces agyában. Neurogenezist azóta
emberben is kimutattak, például a bonyolult téri tájékozódásra
kényszerülő londoni taxisofőrök hippokampuszában. A HVC többek között
az RA-magot idegzi be. A HVC RA-ba projektáló neuronjai éneklés közben
tüzelnek, egy-egy neuron a motívum egyetlen szótagja éneklésének
egyetlen pillanatában. A pillanat szó szerint értendő, mert az
ismétlések során ugyanaz a neuron 10 milliszekundumos pontossággal
tüzel ugyanazon a ponton. A HVC helyi hűtésekor az időbeli mintázat
nem változik, csak lassul. Ez arra utalt, hogy az alapritmust a HVC
nem kívülről kapja, hanem maga generálja, amire most már közvetlen
bizonyíték is van: a motoros parancsmintázatot a HVC saját
neuronjainak kapcsolatrendszere biztosítja egyfajta dominóelv alapján
(Long et al., 2010).
Úgy tűnik tehát, hogy a HVC a saját ének motoros
parancsmintázatát tárolja, azaz a megtanult tutor templát és a saját
ének motoros mintázata térben valamennyire elkülönül, legalábbis
felnőtt madárban. A HVC által beidegzett RA-terület egyes neuronjai
éneklés közben többször tüzelnek. Nem eldöntött, hogy felnőtt madárban
pontosan mit és miként kódol az RA-terület, mivel azonban ez a mag a
vokalizációt közvetlenül biztosító motoros magokat idegzi be (nem
szerepel az 1. ábrán), és ez az utolsó állomás, ahol az ének
teljes parancsmintázatának össze kell állnia, valószínűsíthető, hogy
nem az egyes idegsejtek, hanem sok idegsejt tüzelési mintázata
reprezentálja a motoros parancsmintázatot.
Mivel a felnőtt madár a HVC- vagy RA-területek
lézióját követően nem énekel, a HVC – RA-pályát hagyományosan
„motoros” pályának tartották. Újabb keletű közvetlen bizonyítékok
azonban arra hívják fel a figyelmet, hogy a motoros pálya területei
érzékenyek a saját dallam érzékelésére, tehát a motoros és szenzoros
funkció együtt van jelen.
Éneket tanuló pályák az agyban
A HVC szükséges a tanult ének produkciójához, de nem elégséges a
tanuláshoz. A tanulásban egy másik rendszer, az előagyi anterior pálya
vagy másik nevén a szenzomotoros pálya magjai kritikusak. A
HVC a striatalis X-területet (Area X) idegzi be, az
X-terület a talamikus DLM-maghoz küld axonokat, a DLM az LMAN-magba,
az LMAN-mag pedig egyrészt az X-területbe projektál, bezárva ezzel a
kört, másrészt az RA-magba, visszakapcsolva az anterior pályát a
motoros pályához (1. ábra). Az anterior
előagyi pálya mind kapcsolatrendszerében, mind a részletekben, például
a neurotranszmitter receptorok eloszlásában analóg – talán homológ – a
motoros és beszédtanulásában fontos emlős kéreg – bazális ganglionok –
talamusz – kéreg pályarendszerrel. A madárban azonban ez a rendszer
egyszerűbb – mondjuk inkább, letisztultabb –, mert mintegy
célszerszámként alapvetően egy funkciót szolgál, és ezért könnyebben
tanulmányozható.
Fiatal madár az anterior pálya sérülése után nem
tanul, ám a felnőtt madár változatlanul énekel tovább, ez a rendszer
tehát elsősorban nem a produkcióhoz, hanem a tanuláshoz szükséges. Az
X-területre projektáló LMAN-mag léziója után a fiatal madár tovább
énekel, de nem tanul, az ének túl korán kristályosodik. Érdekes módon
az X-terület léziójának hatása ellentétes, az ének nem kristályosodik,
és a madár öreg fejjel is mintegy gyakorolja a dallamot. A klasszikus
léziós kísérletek eredményeit más, finomabb vizsgálatok (pl. korai
gének expressziója) is megerősítették. Feltételezhető, hogy az
anterior előagyi pálya a bazális ganglion rendszerhez hasonlóan az
előrejelzési hibát (predicition error) méri, és ha a saját ének a
tanult templáttól különbözik, megnöveli a premotoros mag
plaszticitását, a madár módosít a hibás dallamon. Több adat utal arra,
hogy a javítás nem egyszerű próba-szerencse alapon, hanem irányítottan
történik, bár még nincsen konszenzus abban, hogy a folyamat modellje
alapvetően szelekcionista vagy inkább instrukciós. Az sem egyértelmű,
hogy a hibajavítás kizárólag az anterior pályán zajlik-e, mert a saját
ének hallása nem aktiválja ezt a rendszert, a vizsgálatokba ezért
újabban más területeket, elsősorban a bazális ganglionokat is
bevonják.
Az utóbbi néhány év eredményei tovább finomították
a képet. Hagyományosan a két pályarendszert az énektanulás fázisainak
megfelelően a produkcióhoz (motoros pálya), illetve a tanuláshoz
(szenzorimotoros pálya) társították, azonban kiderült, hogy az
érzékelés és a motoros funkció nem válik szét élesen. Az előagyi
anterior pálya sérülése esetén a fiatal madár nem produkál
elődallamokat, és a gügyögés elmaradásának oka alapvetően a motoros
deficit.
Génekről a kultúráig
Ebben a szakaszban néhány érdekes, új eredményről számolok be, amelyek
új utat nyithatnak a madárének-tanulás megértésében, és ötleteket
nyújthatnak a beszéddel foglalkozó kutatók számára.
Venetianer Pál részletesen ismerteti egy gén, a
FoxP-2-gén mutációjának érdekes következményeit az emberi beszédre, az
egerek vokalizációjára. A FoxP-2-gén számos szervben expresszálódik,
kiesése drasztikus következményekkel jár, a mutáció homozigóta
formában letális. Mindez azt valószínűsíti, hogy a FoxP-2 nem
specifikus beszédgén, de többek között a beszédhez szükséges hálózat
megfelelő kialakulásához és működéséhez is szükséges. Hasonló szerepe
lehet
|
|
|
a madarak énektanulásában. A FoxP-2 mRNS-szint
csökkentése az X-területen rontja az énektanulást, a manipulált fiatal
zebrapintyek pontatlanul másolják a tutor dallamát (Haesler et al.,
2007). Az X-területre a ventrikuláris zónából vándorolnak be az
újonnan keletkezett neuronok. Ha ebben a zónában csendesítik le a
FoxP-2-gén működését, a neurogenezis nem szenved mennyiségi zavart, de
a létrejövő idegsejteken kevesebb lesz a dendrittüske, az új neuronok
mintegy butábbak a kontrolállatokhoz képest (Schulz et al., 2010).
A gének expressziós mintázata az éneklő madár
különböző agyterületein ma jól vizsgálható. A génexpressziós elemzések
arra utaltak, hogy az énekléskor, illetve a hallott dallam
észlelésekor aktiválódó gének száma eléri a százat. A vizsgálatoknak
új lendületet ad, hogy publikálták az első énekesmadár genomot (Warren
et al., 2010), amelyet a házityúk és az ember genomjával összevetve
eddig ismeretlen génekre is ráirányul a figyelem. Az előzetes
eredmények alapján úgy tűnik, hogy az énekkel kapcsolatos gének száma
eléri a 800-at, és ezek között szabályozó RNS-molekulákat kódoló gének
is vannak. Érdekes, hogy a szabályozó RNS-ek egy része mikro RNS,
melyek egy bizonyos traszkripciós faktor fehérje megfelelő helyeire
képesek kötődni, zebrapintyben éppúgy, mint emberben. Remény van arra,
hogy a következő évtizedben a zebrapinty éneklésének molekuláris
vizsgálata elvezet a beszéd molekuláris szabályozásának feltárásához.
A látott mozgásmintázat utánzásában fontos szerepe
van a tükörneuron-rendszernek. A tükörneuronok akkor tüzelnek, ha egy
látott mozgást utánozunk, tehát ezek kapcsolják össze a vizuális és
motoros információt, megkönnyítve az utánzásos tanulást. Szerepük
nagyon fontos az empátiában is, például autista gyerekek
tükörneuron-rendszerének aktivitása az átlagosnál alacsonyabb. A
hallott ingerek összevetése a motoros mintázattal a nem topografikus
leképezés miatt bonyolult. Ezt a folyamatot segíti a madarak
énektanulásában és énekfelismerésében aktív tükörneuron-rendszer,
amelyet a HVC X-területre projektáló neuronjai között fedeztek fel
(Prather et al., 2008). A felfedezés jelentősége túlmutat az
énektanulás körén, feltételezhető, hogy a beszédmegértésben,
gyakorlásban is hasonló rendszer működik, így most már emberben is
célzottan kereshető; ezen túl, a tudományos előrehaladást gyorsítja,
hogy a türkörneuron-rendszer kis testű, könnyen manipulálható állatban
vizsgálható.
A hagyományos elképzelés szerint a fiatal madár
tanul, és amit tud, auditoros visszacsatolás nélkül is, változatlan
formában adja elő. Egy olyan modell, amelyben fiatal korban lezárul a
viselkedés és az idegrendszer plaszticitása, nem túl érdekes az ember
számára. A madárkutatók szerencséjére, ma már tudjuk, hogy a madarak
alkalmazkodóképessége felnőtt korban is megmarad. Erre példa a
vizsgálat, amely során az ének megváltoztatására késztettek felnőtt
madarakat (Sober – Brainard, 2009). Japán sirálykák énekét mikrofonon
keresztül egy áramkörrel manipulálták, és a madár a saját énekét egy
fülhallgatón keresztül hallotta vissza. A manipuláció lényege az volt,
hogy az ének négy szótagjának domináns frekvenciáját megemelték, tehát
a madár a saját énekét a vártnál magasabb fekvésben hallotta. Ennek
kiigazítására a sirálykák két héten át minden nap egyre mélyebb hangon
énekeltek.
A felnőtt ének plaszticitását a fiatalokhoz
hasonlóan az anterior előagyi pálya biztosítja, szükség esetén mintegy
felszabadítva a premotoros RA-magot a HVC szigorú kontrollja alól.
Érdekes, hogy a fiatal zebrapintyekben az RA-mag alvás közben is
aktív, és az énekprodukciónak megfelelő mintázatot mutat. Az alvás
előtti mintázathoz képest az RA-neuronok nagyjából fele némileg eltérő
mintázatban tüzel közvetlenül alvás után (Rauske et al., 2010), ami
más vizsgálatokkal együtt arra utal, hogy az alvás közben lezajló
folyamat aktív része a tanulásnak. A tutor énekhez való
hasonlatosságban mérve a fiatal zebrapintyek alvás után azonban
hibásabban énekelnek, mint előző este, tehát az alvás nem mélyíti el a
memórianyomokat. Az alvás utáni rontás mégis fontos a végső dallam
kialakulásában, mert azok a pintyek produkálták végül a legjobb
másolatot, akik a legrosszabbul énekeltek reggelente (Derégnaucourt et
al., 2005)! A pintyek saját éneküket a tanult templáttal hasonlítják
össze gyakorlás közben, és memorizálják saját motoros mintázatukat. A
tanulási folyamatnak egyaránt része a merev memórianyomok képzése és
rugalmas megváltoztatása. Úgy tűnik, az álom nem a memórianyomok
rögzítésében segít, hanem épp ellenkezőleg, az RA-neuronok
felszabadításával, mintegy kreatívvá teszi a rendszert. Reggel a
fiatal madár többféle megálmodott változatot próbál ki, kiválasztja a
legjobb utánzatot, s rögzíti a motoros parancsmintázatot.
Említettem, hogy az izolációban felnevelt
zebrapintyek is énekelnek, a dallam tükrözi a fajra jellemző
szerkezetet, de torz formában és az egyedi variációk nélkül. Mi
történik, ha szegényes és torz izolált éneket játszunk le mintaként a
fiatal hímek első generációjának? Tovább rontják-e az ének minőségét,
és a hiba mértéke generációról generációra növekszik? Érdekes módon
ennek az ellenkezője történik, az izolált éneket megtanuló fiatalok
dallama jobb, mint a templát, és az ének generációról generációra
egyre jobban hasonlít a vad típusra (Fehér et al., 2009). A
genetikailag kódolt fajspecifikus alapmintázatot a fiatalok két
szempontból módosították, egyrészt az abnormálisan hosszú szótagokat
lerövidítették, másrészt új, egyedi motívumokat is beleszőttek az
eredeti szegényes énekbe. Ezzel egyedi, de a természetes populációkra
jobban emlékeztető új kultúra jött létre, amelyet sokan az emberi
kultúra, nyelv jól vizsgálható modelljének tartanak. A párhuzam
azonban nem annyira a nyelv, mint a beszédprodukció evolúciójával
vonható meg, mert gyakorlatilag semmit sem tudunk arról, van-e a
kogníciónak szerepe a zebrapintyek énektanulásában.
A gondolkodó madár
Tyúkeszűnek (birdbrain) nevezni bárkit, semmilyen nyelven nem számít
bóknak még ma sem, pedig már tudjuk, hogy egyes madárfajok tudatosan
készítenek eszközöket, van számosságfogalmuk, epizodikus memóriát
képeznek, rendelkeznek elmeteóriával (theory of mind), és képesek
mentális időutazásra, azaz tudatosan felkészülnek arra, hogy a
következő nap hol és mit fognak csinálni. Az apró énekesmadarak is
okosak, a mi laboratóriumunkban a zebrapintyek gyorsan megtanulják,
hogy egy libikókára ugorva társukat fel tudják emelni a másként nem
elérhető finom falatokhoz.
Mivel gondolkodnak a madarak? A hagyományos képzet
szerint az emlősagy olyan madáragy, amelyre kérget húzott az evolúció,
tehát az emlősök agya több és minőségileg jobb a madarakénál. Mivel a
neokortex nélkülözhetetlen a tanuláshoz és gondolkodáshoz, a kérget
nélkülöző agy csak egyszerű reflextevékenységeket képes szabályozni.
Ez az elképzelés ma már nem tartható, mert sok, független bizonyíték
alapján tudjuk: a striátumnak vélt régiók jelentős része valójában
pallidum, és megfelel az emlősök neokortexének (ABNC, 2005). Az
emlősagy tehát nem minőségében, hanem szerkezetében más.
Mivel a madarak kognitív képességei vetekednek az
emlősökével, meglepő, hogy az énekesmadarak vokalizációjában a
kogníció meggyőző lenyomatát még nem sikerült kimutatni. Az
énekesmadarak dalai elsősorban az egyed minőségét hirdetik, és
szexuális szelekció alatt állnak. Beteg madár gyengén énekel,
táplálékínségben fölnevelt madarak HVC- és RA-magja kicsi, énekük
rossz utánzat, s ezeket a hímeket a tojók gyengébb minőségűnek
értékelik (Nowicki et al., 2002). Sok énekesmadár a körülményeknek
megfelelően változtatja vokalizációját. Zajos városokban a feketerigók
hangosabban énekelnek vidéki társaiknál, és daluk komplexitása is
fokozódik, ha sok a vetélytárs a környéken. A plaszticitás tehát
részben válasz lehet a fizikai környezet korlátaira, de elsősorban az
egyed belső állapotát tükrözi, a madárének nem tekinthető szemantikus
jelzésnek. Ez azért lehet meglepő, mert néhány madár vész- vagy
csődítő kiáltásának van jelentéstartalma. A házikakas másként kiált,
ha földi vagy légi ragadozót észlel. A feketefejű cinege a különböző
ragadozók láttán ugyanazt a csődítő kiáltást hallatja, de a kiáltás
végén elhangzó „dí” szótag száma arányos a ragadozó méretével. Az
énekkel szemben tehát a kiáltásnak sok esetben van jelentése, de hogy
a nem tanult kiáltást a madarak öröklött reflexek vagy tapasztalataik
alapján módosítják, nem tudjuk. A tapasztalat szerepére utal, hogy egy
másik faj, a kanadai csuszka is megfelelő módon reagál, „érti” a
feketefejű cinege kiáltásának jelentését. Jelenleg annyit mondhatunk,
hogy a madarak énektanulásában nem, de a vokalizáció alkalmazásában
kimutathatóak a kogníció jelei. Nem tudható, hogy a gazdag
repertoárral rendelkező trópusi fajok alaposabb vizsgálata
változtatna-e ezen a képen.
Az énekesmadarakon kívül a kolibrik és a papagájok
is tanulják a vokalizációt. A papagájok, legalábbis egy afrikai
szürkepapagáj, a nemrég elhunyt Alex képes volt összekötni a
vokalizációt magas szintű kognitív folyamatokkal. Alex angol nyelven
kimondta tárgyak nevét, színét, anyagát, megszámolta a tárgyakat
hatig, értette és használta a kisebb–nagyobb, azonos–különböző stb.
relációt, és képes volt ezeket egymásba ágyazva használni. Meg tudta
például mondani, hogy hány piros kocka van egy tálcán, amelyre
különböző színű kockákat és golyókat helyeztek. Meg tudta mondani,
hogy mi a különbség két tárgy között, például a színük vagy az anyaguk
más. Alex valószínűleg gyakorolta az emberi gondolkodás legnagyobb
vívmányait, a hazudozást és a semmi fogalmát. A tálcán lévő két piros,
három kék és négy zöld kocka alapján válaszolnia kellett arra a
kérdésre: milyen szín három?. A papagáj nem akart dolgozni, helyette
csikizést kért, és a kérdésre többször azzal válaszolt, hogy öt. Végül
Irene Pepperberg azt mondta: Na jól van, milyen szín öt?, mire
Alex azt mondta: Semmi.
Alex adekvát módon használta az igéket és
főneveket, „beszéde” azonban nélkülözte az emberi nyelv sajátosságait.
Mindez arra utal, hogy a madarak éneke jó modell az emberi beszéd, de
nem az emberi nyelv biológiájának megértéséhez. Ám ahogy a közelmúlt,
a közeljövő is tartogat meglepetéseket.
Kulcsszavak: madárének, tanulás, madarak, nyelv, kogníció
IRODALOM
ABNC – Avian Brain Nomenclature Consortium
(2005): Avian Brains and a New Understanding of Vertebrate Brain
Evolution. Nature Reviews Neuroscience. 6, 151–159. •
WEBCÍM >
Bolhuis, Johan J. – Okanoya, K. – Scharff,
C. (2010): Twitter Evolution: Converging Mechanisms in Birdsong and
Human Speech. Nature Reviews Neuroscience. 11, 747–759. •
WEBCÍM >
Derégnaucourt, Sébastien – Mitra, P. P. –
Fehér O. – Pytte, C. – Tchernichovski, O. (2005): How Sleep Affects
the Developmental Learning of Bird Song. Nature. 433, 710–716. DOI:
10.1038/nature03275
Fehér Olga – Wang, H. – Saar, S. – Mitra,
P. – Tchernichovski, O. (2009): De Novo Establishment of Wild-Type
Song Culture in the Zebra Finch. Nature. 459, 564–568.
Haesler, Sebastian – Rochefort, C. –
Georgi, B. – Licznerski, P. – Osten, P. – Scharff, C. (2007):
Incomplete and Inaccurate Vocal Imitation After Knockdown of Foxp2 in
Songbird Basal Ganglia Nucleus Area X. Plos Biology, 5, E321 •
WEBCÍM >
Long, Michael A. - Jin, D. Z. – Fee, M. S.
(2010): Support for a Synaptic Chain Model of Neuronal Sequence
Generation. Nature. 468, 394–399. •
WEBCÍM >
Mátyás Ferenc – Sreenivasan, V. – Marbach
F. – Wacongne, C. – Barsy B. – Mateo, C. – Aronoff, R. – Petersen, C.
C. H. (2010): Motor Control by Sensory Cortex. Science. 26, 1240–1243.
•
WEBCÍM >
Mooney, Richard (2009): Neurobiology of
Song Learning. Current Opinion in Neurobiology. 19, 654–660.
Nowicki, Stephen – Searcy, W. A. – Peters,
S. (2002): Quality of Song Learning Affects Female Response to Male
Bird Song. Proceedings of the Royal Society London B. 269, 1949–1954.
•
WEBCÍM >
Pepperberg, I. (2002): Cognitive and
Communicative Abilities of Grey Parrots (Psittacus Erithacus). Current
Directions in Psychological Science. 11, 83–87. •
WEBCÍM >
Prather Jonathan F. – Peters S. – Nowicki
S. – Mooney R. (2008): Precise Auditory-Vocal Mirroring in Neurons for
Learned Vocal Communication. Nature. 451, 305–10.
doi:10.1038/nature06492
Rauske, Peter L. – Chi, Z. Y. – Dave, A.
S. – Margoliash, D. (2010): Neuronal Stability and Drift Across
Periods of Sleep: Premotor Activity Patterns in a Vocal Control
Nucleus of Adult Zebra Finches. The Journal of Neuroscience. 30,
2783–2794. •
WEBCÍM >
Riede, Tobias – Fisher, J.H. – Goller, F.
(2010): Sexual Dimorphism of the Zebra Finch Syrinx Indicates
Adaptation for High Fundamental Frequencies in Males. Plos ONE. 5, 6,
E11368 •
WEBCÍM >
Schrott Anikó – Kabai Péter (2008): ABCD:
a Functional Database for the Avian Brain. Journal of Neuroscience
Methods. 167, 393-395.
Schulz, Steffen – Haesler, S. – Scharff,
C. – Rochefort, S. (2010): Knock-Down of Foxp2 Alters Spine Density in
Area X of the Zebra Finch. Genes Brain and Behaviour, 9, 732–740. DOI:
10.1111/j.1601-183X. 2010.00607.x
Sober, Samuel J. – Brainard, Michael S.
(2009): Adult Birdsong Is Actively Maintained by Error Correction.
Nature Neuroscience. 12, 927–931. •
WEBCÍM >
Warren, W. C. et al. (2010): The Genome of
a Songbird. Nature. 464, 757–762. •
WEBCÍM >
LÁBJEGYZETEK
1 Színes ábrák, letölthető
források:
WEBCÍM >
|
<
2 Az énektanulás idegi
hátterének részletes ismertetése meghaladja a dolgozat kereteit. Csak
a fontosabb területeket említem, és ezeknek is csak a rövidítését. A
madarak agyterületeit és a közöttük lévő ismert kapcsolatok
böngészését Schrott Anikó és Kabai Péter (2008) adatbázisa könnyíti
meg:
WEBCÍM >
|
<
3 A madáragy-anatómusok
között nincs teljes konszenzus abban, hogy az egyes magok miként
feleltethetők meg az emlős agykéreg területeinek, az sem világos, hogy
a megfelelés evolúciós értelemben analógiának vagy homológiának
tekinthető-e (ABNC, 2005).
<
4 A HVC jelenleg a „magas
vokális központ” (High Vocal Center) rövidítése. Az anatómiában
szokatlan elnevezésnek szokatlan története van. A HVC-magot Fernando
Nottebohm írta le a hiperstriátum részeként (hyperstriatum ventrale
caudale). Később kiderült, hogy a mag valójában a neostriátumban van,
de a rövidítés – akkor még kis c-vel (HVc) annyira elterjedt, hogy nem
változtatták meg, inkább a rövidítésnek adtak új nevet. Pár éve egy
konzorcium (ABNC 2005) a madár telencephalon területeinek zömét
átnevezte, az új terminológiában neostriatum sincs, helyette
nidopalliumot mondunk.
<
|
|