|
keresztül az adott molekula végét meg lehet
ragadni, és a molekulára erővel lehet hatni. A kifejtett erők igen
alacsonyak, akár femtonewton nagyságrendűek lehetnek (fN = 10-15 N). A
mágneses csipesz használatakor különösen nagy jelentőségű, hogy a
külső mágnes elforgatásával a manipulált mikrogyöngy is elfordul. Ha a
megragadott molekula vége szilárdan rögzül a mikrogyöngy, illetve a
szubsztrát felszínéhez, akkor a molekulára torziós erőt fejthetünk ki.
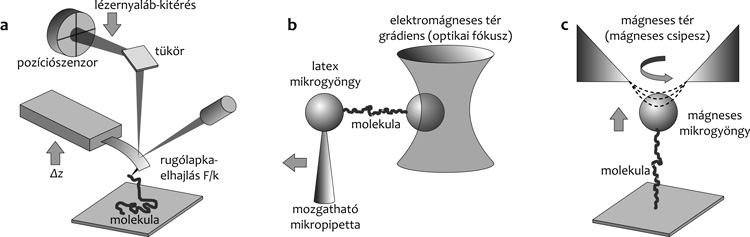
4. Molekulák rögzítése • Annak érdekében, hogy
egyedi biomolekulákat mechanikailag manipuláljunk, a végeit megfelelő
módszerrel meg kell fognunk. A lézercsipeszben a molekula végeit
különböző felszíni reaktív csoportokat hordozó mikrogyöngyökhöz
kapcsoljuk. Ezután az aktivált mikrogyöngy fogantyúként viselkedik,
mellyel a vizsgált molekulát manipulálni tudjuk. Az AFM-ben a molekula
a tű és a felszín között feszül ki, melyek mindegyike kémiailag
aktiválható. Az egyedi DNS-molekulák manipulálására alkalmas technikák
széles skálája megteremtette annak lehetőségét is, hogy magát a
DNS-molekulát mint fogantyút alkalmazzuk. Az utóbbi években különösen
nagy érdeklődés irányul a kémiailag funkcionalizált szén nanocsövekre,
melyek az AFM-ben alkalmazhatók fogantyúként.
Mire lehet következtetni
egy molekula manipulálásából?
Egyedi molekulák manipulációjából számos tulajdonságra, állapotra és
folyamatra lehet következtetni: rugalmasság, szerkezeti állapotok,
intramolekuláris kölcsönhatások, erőgeneráló folyamatok.
1. Mért paraméterek • Az egymolekula manipulációs
kísérletekben nyert adatok a mechanikai erő, illetve a molekula
felfüggesztési pontjai közötti távolság. Mindkét paraméter időbeli
változásait is követhetjük nagy felbontással. Az erő a molekulában
fejlődik ki a mechanikai perturbáció hatására. A felfüggesztési pontok
közötti távolság láncszerű molekula esetében többnyire a molekula két
végpontja közötti távolság.
2. Mechanikai kalibráció • Az egyedi molekula
manipulációs eszközök parányi erők (fN–nN tartomány) mérésére alkalmas
erőátalakító berendezések. Hogy a vizsgált molekulában kifejlődő erőt
pontosan megmérhessük, ismernünk kell az erőtranszducer (például
lézercsipesz vagy AFM-rugólapka) rugóállandóját. A rugóállandót
kalibrációs protokollok segítségével mérjük meg, melyek alapja ismert
nagyságú (például viszkózus, termikus) erőhatásra fellépő elmozdulás
vagy elhajlás megmérése.
3. Molekularugalmasság • Annak érdekében, hogy egy
fehérjelánc rugalmasságát megmérjük, a molekulát végeinél fogva meg
kell nyújtanunk. Lézercsipesz esetében például a molekula mindkét
végére egy-egy mikrogyöngy kapcsolódik. Az egyik mikrogyöngyöt a
lézercsipesszel, a másikat egy mozgatható mikropipettával fogjuk meg
(1. b ábra). A molekulát a mikropipetta lézercsipesztől történő
eltávolításával nyújtjuk meg, majd a mikropipetta lézercsipesz felé
történő visszamozgatásával relaxáljuk a molekulát. A
nyújtás-visszaengedés ciklus során mérjük vagy kiszámoljuk a molekula
két vége közötti távolságot és az ehhez tartozó rugalmas erőt. Más
módszerek, például az AFM esetében (1.a ábra) a molekula végeit
a rugólapka tűjével, illetve a szubsztrát felszín segítségével
ragadjuk meg. Hosszú biopolimér-láncok esetében a rugalmas
erő-molekulahossz függvény többnyire nemlineáris összefüggést mutat,
utalva arra, hogy a rugalmassági mechanizmus elsősorban entropikus, és
nem a merev testek rugalmassági törvényeit követi. Egy entropikus
polimer összehúzódik, mert benne termikusan gerjesztett hajlító
mozgások lépnek fel, melyek növelik a lánc konfigurációs entrópiáját.
A konfigurációs entrópia csökkenthető a lánc megnyújtásával, azaz a
lánc végeinek egymástól való távolításával. Ehhez külső mechanikai
erőre van szükség. Ahogy a lánc vég–vég távolsága közelíti a
kontúrhosszt, az erő aszimptotikusan nő. A molekula rugalmassági
mechanizmusainak pontos megértése érdekében a kísérleti erő-megnyúlás
görbére elméleti függvényeket illesztünk, így kaphatjuk meg az
entropikus polimérlánc hajlítási rugalmasságát leíró úgynevezett
perzisztenciahosszt.
4. Erő hatására fellépő szerkezetváltozások •
Mechanikai erő hatására nem csupán reverzibilis, rugalmas alakváltozás
lép fel a molekulában, hanem szerkezetváltozások is. Éppen a
szerkezetváltozások detektálása és jellemzése vezet el az adott
molekula tulajdonságainak és működésének pontos megismeréshez. Az
alábbiakban konkrét példán, a titin óriás izomfehérjén mutatjuk be az
erővezérelt intramolekuláris szerkezetváltozásokat.
A titin volt a legelső fehérje, amely egyedi
molekulamanipulációs technikákkal manipulálásra, megnyújtásra került
(Rief et al., 1997; Tskhovrebova et al. 1997; Kellermayer et al.
1997). A harántcsíkolt izom összehúzódásán kívül rugalmas
tulajdonságokkal is rendelkezik, melyekért elsősorban épp ez a
különleges, hatalmas méretű filamentális fehérje a felelős. Az
izomrostban megnyújtásra fejlődő erőt passzív izomerőnek nevezzük, és
fontos szerepe van az izomszarkomer szerkezeti integritásának
fenntartásában, illetve a nyújtási erő megszűnte után az izom nyugalmi
hosszának helyreállításában. A titinről ma már tudjuk, hogy mérete, a
szarkoméren belüli elhelyezkedése és egyéb fehérjékkel való
kölcsönhatásai miatt egyfajta organizáló, rendező szerepet játszik az
izomban. A titin közel 4 MDa tömegű, láncszerű fehérje, amelyet
mintegy 300, sorbakapcsolt globuláris fehérjedomén és a közöttük
itt-ott elhelyezkedő egyedi szekvencia épít fel. A globuláris domének
hét, antiparallel irányban futó béta-láncból épülnek fel, amelyek,
mint a hordó dongái, egy hordó alakú szerkezetet építenek fel.
Érdekes, hogy a fehérjeszerkezet alapján a titin globuláris doménjei
hasonlítanak bizonyos extracelluláris fehérjék, az immunglobulinok és
a fibronektin doménjeihez.
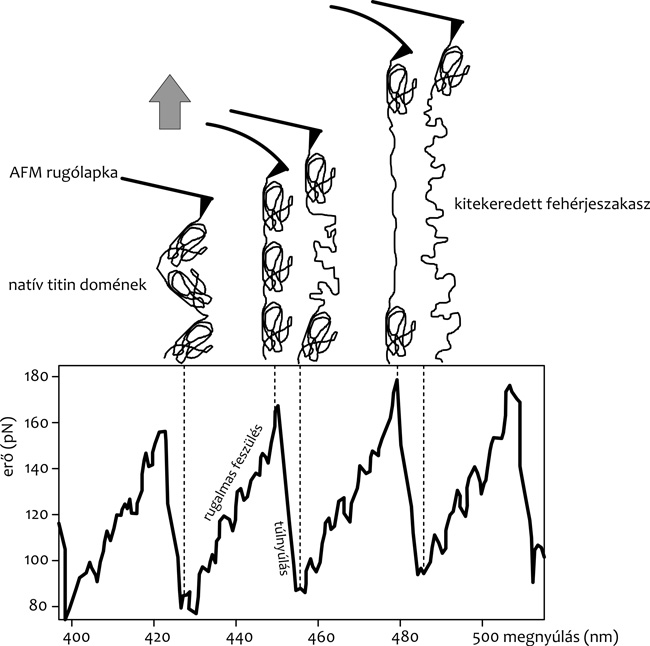
Ha egy titinmolekulát végeinél fogva megnyújtunk,
benne rugalmas erő ébred (2. ábra). A titin rugalmassága jól
jellemezhető entropukis polmérmodellek segítségével. Ha azonban az erő
meghalad egy bizonyos értéket, az erőgörbe hirtelen eltér a rugalmas
modell szerint várható lefutástól, és a molekula túlnyúlik, mintegy
megfolyik. A titin túlnyúlásának oka a molekulát felépítő globuláris
domének erő hatására fellépő kitekeredése. Az egymást követő egyedi
doménkitekeredési események fűrészfog alakú erőgörbét eredményeznek.
5. Erőspektrum • A bonyolult szerkezeti átmenetek
sorozatát tartalmazó erőgörbét erőspektrumnak nevezzük. Az
erőspektrumban észlelt pillanatnyi erőváltozások intra- vagy – a
kísérleti geometriától függően – intermolekuláris kölcsönhatások
mechanikailag vezérelt felszakadására vezethetők vissza. A
kölcsönhatások ennek megfelelően lehetnek például
fehérje térszerkezetet össszetartó kötések, antigén/antitest kötések,
vagy ligandum/kötőhely kölcsönhatások (például
biotin/sztreptravidin). Ha egy láncszerű molekulában erő hatására
felszakadó kölcsönhatások diszkrét, lépésszerű megnyúlást okoznak, az
erőspektrumban fűrészfogak jelennek meg (2. ábra).
Egy molekula vagy molekulakomplex szerkezetét a benne fellépő
kölcsönhatások egyensúlya és így az asszociációs és disszociációs
reakciók relatív sebességei határozzák meg. Könnyű belátni ezért, hogy
az erő hatására fellépő szerkezetváltozás attól is függ, hogy milyen
sebességgel terheljük a molekulát. Az erőspektrum terhelési sebesség
függvényében történő analízisét dinamikus erőspektroszkópiának
nevezzük. A dinamikus erőspektroszkópia segítségével gyakran nehezen
hozzáférhető termodinamikai és kinetikai paraméterek válnak
megmérhetővé.
Perspektívák
Az utóbbi években az egyedi molekulák vizualizálásában és mechanikai
manipulálásában látványos fejlődésnek lehettünk tanúi. Fontos
felfedezések történtek, és a módszerek egy része széles körben
alkalmazásra került. A tudományterület fejlődési irányát nehéz
megjósolni. Valószínű azonban, hogy további technikai fejlődés
(például érzékenyebb szenzorok, egyre gyorsabb adatvétel) és különböző
egymolekula módszerek kombinációi meghatározó tényezők lesznek.
Bizonyos módszerkombinációk már jelenleg is léteznek. A képalkotó AFM
ötvözése molekuláris erőspektroszkópiával egyedi információval szolgál
a lokális szerkezetről, dinamikáról, mechanikáról. Egymolekula
manipulációs módszerek kombinációja fluoreszcencia mikroszkópos és
spektroszkópos technikákkal széles térbeli és időskálán történő
méréseket tesz lehetővé. Egy különösen látványos kísérleti
elrendezésben a lézercsipesz és egyedi molekula fluoreszcencia
kombinálásával sikerült a miozin mechanikai működését és enzimatikus
funkcióját egyszerre vizsgálni (Funatsu et al., 1995). Számos törekvés
történik az AFM és fluoreszcencia mikroszkópia kombinálására
(Kellermayer et al., 2006). Ezek a kombinációk gyakran szekvenciálisak
(vagyis időben egymást követik a mérések) a bonyolult geometria, az
igen eltérő adatvételi sebességek és az egyedi fluorofór detektálására
elégtelen érzékenység miatt. Egyedi, mechanikailag manipulált
fehérjemolekulák fluoreszcenciájának követésével lehetőség nyílik
nagyfelbontású intramolekuláris szerkezeti és dinamikai vizsgálatokra.
Figyelembe véve, hogy keveset tudunk a mechanikailag perturbált
molekulák nagyfelbontású pillanatnyi szerkezetéről, ezek a
fejlesztések és törekvések igen nagy jelentőségűek. A nemrégiben
kifejlesztett szuperrezolúciós mikroszkópos technikák újabb kapukat
nyitnak egyedi molekulák viselkedésének pontosabb megismerésére (Huang
et al., 2008; Lippincott-Schwartz – Manley 2009). Végső soron az sem
reménytelen, hogy megfelelő kémiai, genetikai és mechanikai
manipulációk segítségével olyan erőérzékelő fluoreszcens molekulák
fejleszthetők, amelyek mintegy láthatóvá teszik az élő sejtben
állandóan működő, eddig alig ismert molekuláris erőhatásokat, útnak
indítva ezáltal egy újabb, már a laboratóriumaink bejáratánál
kopogtató tudományterületet, a mechanobiológiát (Lim et al., 2010).
Kulcsszavak: lézercsipesz, atomerő-mikroszkóp, nanomechanika,
fluoreszcencia, titin, molekuláris rugalmasság, fehérjegombolyodás
IRODALOM
Bustamante, C. – Macosko, J. C. – Wuite, G
J. (2000): Grabbing the Cat by the Tail: Manipulating Molecules One by
One. Nature Reviews Molecular Cell Biology. 1, 2, 1 November, 130–136.
DOI:10.1038/ 35040072.
Dickson, R. M. – Cubitt, A. B. – Tsien, R.
Y. – Moerner, W. E. (1997): On/Off Blinking and Switching Behaviour of
Single Molecules of Green Fluorescent Protein. Nature. 388, 6640, 24
July, 355–358. DOI:10.1038/41048.
Funatsu, T. – Harada, Y. – Tokunaga, M. –
Saito, K. – Yanagida, T. (1995): Imaging of Single Fluorescent
Molecules and Individual ATP Turnovers by Single Myosin Molecules in
Aqueous Solution. Nature. 374, 555–559.
Huang, Bo – Jones, S. A. – Brandenburg, B.
– Zhuang, X. (2008): Whole-Cell 3D STORM Reveals Interactions between
Cellular Structures with Nanometer-Scale Resolution. Nature Methods.
5,12, 1 December, 1047–1052. DOI:10.1038/nmeth.1274.
Kellermayer Miklós S. Z. – Smith, S. B. –
Granzier, H. L. – Bustamante, C. (1997): Folding-Unfolding Transitions
in Single Titin Molecules Characterized with Laser Tweezers. Science.
(New York, NY) 276,5 315, 16 May, 1112–1116.
Kellermayer Miklós S. Z. (2005):
Visualizing and Manipulating Individual Protein Molecules.
Physiological Measurement. 26, 4, 1 August, R119–R153.
DOI:10.1088/0967-3334/26/4/R02.
Kellermayer Miklós S. Z. – Karsai Á. –
Kengyel A. – Nagy A. – Bianco, P. – Huber T. – Kulcsár Á. – Niedetzky
Cs. – Proksch, R. – Grama L. (2006): Spatially and Temporally
Synchronized Atomic Force and Total Internal Reflection Fluorescence
Microscopy for Imaging and Manipulating Cells and Biomolecules.
Biophysical Journal. 91,7, 1 October, 2665–2677. DOI:10.1529/biophysj.
106.085456.
Lim, Chwee Teck – Bershadsky, A. – Sheetz,
M. P. (2010): Mechanobiology. Journal of the Royal Society, Interface
/ the Royal Society. 7 Suppl 3, 6 June, S291–S293.
DOI:10.1098/rsif.2010.0150.focus
Lippincott-Schwartz, Jennifer – Manley,
Suliana (2009): Putting Super-Resolution Fluorescence Microscopy to
Work. Nature Methods. 6, 1, 21–23. DOI:10.1038/nmeth.f.233.
Rief, M. – Gautel, M. – Oesterhelt, F. –
Fernandez, J. M. – Gaub, H. E. (1997): Reversible Unfolding of
Individual Titin Immunoglobulin Domains by AFM. Science. (New York,
NY) 276, 5315, 1109–1112.
Tskhovrebova, L. – Trinick, J. – Sleep, J.
A. – Simmons, R. M. (1997): Elasticity and Unfolding of Single
Molecules of the Giant Muscle Protein Titin. Nature. 387, 6630, 15
May, 308–312. DOI:10.1038/387308a0.
Wen, J. – Lancaster, L. – Hodges, C. –
Zeri, A. –Yoshimura, S. – Noller, H. – Bustamante, C. – Tinoco, I.
(2008): Following Translation by Single Ribosomes One Codon at a Time.
Nature. 452, 7187, 9 March, 598–603. DOI:10.1038/nature06716.
|