|
A rendelkezésre álló kísérleti eredményekre
alapozva modellt dolgoztunk ki a HAP2-sapka működésének molekuláris
mechanizmusára nézve, amely magyarázatot nyújt arra, hogy a
HAP2-komplexum miként képes a filamentumok végéhez erősen kötődőve
mégiscsak lehetővé tenni, hogy a centrális csatornán át a
citoplazmából érkező flagellin alegységek alája beépülhessenek
(Vonderviszt et al., 1998). Elképzeléseink szerint a HAP2-sapka
működésében a sapka és a filamentum nemkompatibilis szimmetriája és az
alegységek rendezetlen terminális régióiban a kötődés során
bekövetkező, rendezetlen–rendezett állapotok közötti szerkezeti
átmenetek játszanak meghatározó szerepet.
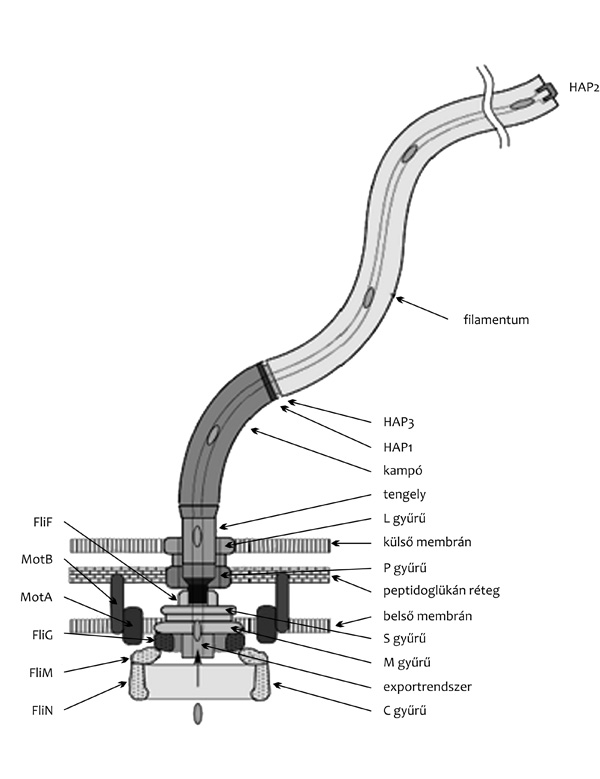
A bakteriális flagellumok axiális fehérjéi egy
speciális exportapparátus segítségével, a filamentumok centrális
csatornáján keresztül jutnak el beépülési helyükre, a filamentumok
végére (Minamino et al., 2008). A flagellumok tövénél a citoplazma
felőli oldalon található exportrendszer képes felismerni az
exportálandó flagelláris fehérjéket, és energiaforrásként a belső
membrán két oldala közötti protonkoncentráció-különbséget kihasználva
kitekeredett formában bejuttatni őket a filamentumok belső
csatornájába. Mindezidáig nem ismert, hogy mi az a jel, szerkezeti
vagy szekvenciális sajátosság, amelynek alapján a flagellumspecifikus
exportapparátus felismeri az exportálandó fehérjéket. Szekvenciális
szinten semmiféle olyan, az axiális fehérjék mindegyikében fellelhető
közös sajátosságot sem sikerült kimutatni, amely az exportapparátus
számára azonosító jelként szolgálhatna. Nincs lehasításra kerülő
szignálpeptid, sem közös szignálszekvencia. Mindez arra utal, hogy az
exportapparátus által felismert közös sajátosság valamilyen magasabb
szerkezeti szinten keresendő.
Különféle génsebészetileg előállított, N-terminális
régiójukban csonkított variánsok exportképességének vizsgálata révén
sikerült lokalizálnunk a flagellin exportszignálját (Végh et al.,
2006; Gál et al., 2006). Megállapítottuk, hogy a flagellin
aminosavszekvenciájának 26-47 szegmense hordozza a felismerési jelet
az exportapparátus számára. Ez az erősen konzerválódott részlet a
flagellin rendezetlen N-terminális régiójában található, amely a
predikciós eljárások szerint jelentős α-hélix-képző potenciállal
rendelkezik. Vajon miként képes egy rendezetlen szegmens felismerési
jelként funkcionálni? Hipotézisünk szerint az exportrendszer felismerő
helye az exportszubsztárok azonosítását végző fehérje alegységen egy
szűk vájatban található, amely csak egy kitekeredett flexibilis
polipeptid szegemens befogadására képes. Az exportszignál a felismerés
során alfa-helikális szerkezetbe rendeződve indítja el a transzlokáció
folyamatát. Hasonló mechanizmus figyelhető meg a mitokondriális
exportrendszer működése során.
A terminális régiók rendezetlensége és azok
helikális stabilizálódási képessége az axiális fehérjék egyedüli közös
szerkezeti sajátossága. Eddig csak néhány flagelláris fehérjénél
történt meg az exportszignál lokalizálása, de azt minden esetben az
N-terminális rendezetlen régió tartalmazza. Mindez a felismerési
folyamat során alátámasztja a helikális rendeződés általános
mechanizmusát.
Megfigyeltük, hogy ha flagellintermelésre képtelen,
ezért csak a kampó részig kiépült exportcsatornát tartalmazó mutáns
szalmonella-gazdasejteket használunk, akkor a flagellin
exportszignálját idegen fehérjékhez kapcsolva azok hatékonyan
kijuttathatók a sejtből a flagelláris exportrendszer segítségével
(Dobó et al., 2010). Ez a felismerés megnyitotta az utat a flagelláris
export célzott felhasználása előtt. Az idegen fehérjék génmódosított
baktériumokkal történő termeltetése napjaink biotechnológiai
gyakorlatának egyik fontos eljárása. Ma már számos enzimet,
hatóanyagot, gyógyszeralapanyagot állítanak elő ilyen módon, azonban
gyakori problémaként jelentkezik, hogy a nagy mennyiségben
termeltetett fehérjetermék rendszerint oldhatatlan formában – ún.
inklúziós testeket képezve – kicsapódik a baktérium belsejében. A
flagelláris exportrendszer kiváló lehetőséget kínál a baktériumokban
nagy mennyiségben termeltetett idegen fehérjék sejtből való hatékony
kijuttatására. Az MTA Enzimológiai Intézetével közösen végzett
fejlesztőmunkánk eredményeként egy olyan bakteriális expressziós
rendszert hoztunk létre (Vonderviszt et al., 2012), amely a széleskörű
biotechnológiai hasznosíthatóság érdekében megkönnyíti a szekretált
fehérje tisztítását és működőképes formában történő előállítását. Az
exportszignál és a termeltetni kívánt fehérje közé beépítettük az
enterokináz enzim specifikus hasítóhelyét, továbbá a fúziós
konstrukció N-terminális végére a tisztítást megkönnyítő polihisztidin
toldalékot tettünk, ami gyakorlatilag egy lépésben lehetővé teszi a
felülúszóból való hatékony izolálást és tisztítást. Ezt követően a
fehérje megfelelő működését esetlegesen zavaró felesleges részek
proteolitikus emésztéssel való szelektív eltávolítása könnyen
megvalósítható. Eljárásunkra szabadalmi védettséget szereztünk.
A szekrécióval összekapcsolt fehérjeexpresszió
számos előnnyel kecsegtet: nincs szükség a sejtek feltárására, ezért a
tisztítandó minta egyéb fehérjékkel, nukleinsavakkal vagy lipidekkel
kevéssé szennyezett. Az eljárás különösen előnyösnek tűnik olyan
fehérjék esetében, amelyek a sejten belül könnyen degradálódnak, vagy
oldhatatlan csapadékot képeznek. Az exportrendszer a flagelláris
fehérjéket részlegesen kitekeredett formában juttatja ki a
filamentumok szűk belső csatornáján keresztül. Feltehetőleg az
exportszignálhoz kapcsolt fehérjék is hasonlóan exportálódnak, azonban
a külső médiumban az aggregációt elkerülve jó eséllyel képesek
felvenni natív működőképes térszerkezetüket.
Flagellinalapú építőelemek
filamentáris nanoszerkezetek létrehozására
A flagellin fehérje képes az önszerveződésre. Vajon miként
ruházhatnánk fel más fehérjéket is az önszerveződés képességével, hogy
belőlük előnyös tulajdonságokkal rendelkező filamentáris
nanoszerkezeteket építhessünk. Munkánk során olyan fúziós fehérjéket
kívánunk létrehozni, amelyekben különféle fehérjéket kapcsolunk össze
a polimerizációra képes flagellin fehérjével, oly módon, hogy mindkét
partner funkcionális tulajdonságai megmaradjanak.
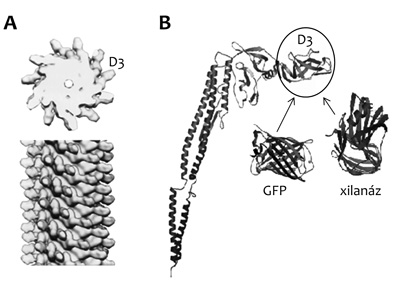
Röntgendiffrakciós és elektronmikroszkópiás
vizsgálatok révén meghatároztuk a flagelláris filamentumok atomi
felbontású térszerkezetét (2/A ábra). A
flagellin alegységek erősen konzerválódott terminális régiói szorosan
egymáshoz kapcsolódva vesznek részt a filamentumépítésben, a
filamentumok belső magjának kialakításában, míg a polipeptidlánc
variábilis középső része alkotja a filamentumok felületén elhelyezkedő
D3-domént, ami a szomszédos alegységekkel nincs kontaktusban.
Vizsgálataink megmutatták, hogy a D3-domén a filamentáris szerkezet
megzavarása eltávolítható, átalakítható, vagy helyére idegen fehérjék
beépíthetők (Muskotál et al., 2010).
A xilanáz enzimet a D3-domén helyére megfelelő
összekötő szegmensek alkalmazásával beültetve a közelmúltban
elkészítettük a polimerizációra képes enzimek prototípusát (2/B
ábra) (Szabó et al., 2011). A flagellin-xilanáz fúziós fehérje
megfelelő körülmények között képes volt filamentumokat formálni,
amelyek xilánbontó katalitikus aktivitást mutattak. Hasonló módon más
enzimek flagellinnel való fúziós konstrukciói is elkészíthetők. A
polimerizációra képes enzimekből akár több ezer katalitikus egységet
hordozó, csatolt reakciók irányítására képes multienzim
nanoszerkezetek építhetők számos alkalmazási lehetőséget kínálva a
biokonverziós technológiákban.
Nemrégiben sikeresen beépítettük a flagellin
belsejébe a zöld fluoreszcens GFP-fehérjét is (Klein et al., 2012). A
GFP-t hordozó flagellin variánsból intenzíven fluoreszkáló
filamentumok építhetők. Jelenleg flagellinalapú kötőfehérjék
előállításán dolgozunk, amelyek képesek célmolekulák specifikus
felismerésére és megkötésére. A flagellinalapú kötőfehérjékből olyan
kontrollált méretű filamentáris nanoszerkezeteket kívánunk építeni,
amelyek felületükön specifikus kötőhelyek ezreit tartalmazzák,
rendkívül nagy kötőhelysűrűséget és az adott célmolekula hatékony
megkötését eredményezve. Filamentáris receptorainkat bioszenzorok
felületére rögzítve specifikus molekulafelismerési tualjdonságokkal
rendelkező fehérjeréteget hozhatunk létre. Az MTA TTK MFA
Nanoszenzorikai Laboratóriumával együttműködésben ilyen érzékelő
réteget alkalmazó, nagy érzékenységű hullámvezető optikai módszerek
kifejlesztésén dolgozunk, amelyek reményeink szerint széles körűen
alkalmazhatóak lesznek a környezeti monitorozásban, az
élelmiszerbiztonsági vizsgálatokban, valamint az orvosi
diagnosztikában.
Kulcsszavak: bakteriális flagellum, flagellin, önszerveződés,
flagelláris export, bakteriális fehérjetermelés, filamentáris
nanoszerkezet
IRODALOM
Dobó József – Varga J. – Sajó R. et al.
(2010): Application of a Short, Disordered N-Terminal Flagellin
Segment, a Fully Functional Flagellar Type III Export Signal, to
Expression of Secreted Proteins. Applied Environmental Microbiology.
76, 891–899. DOI: 10.1128/AEM.00858-09 •
WEBCÍM >
Gál Péter – Végh B. M. – Závodszky P. –
Vonderviszt F. (2006): Export Signals. Nature Biotechn. 24, 900–1. •
WEBCÍM >
Klein Ágnes – Tóth B. – Jankovics H. et
al. (2012): A Polymerizable GFP Variant. Protein Engineering Design
and Selection. DOI: 10.1093/Protein/Gzs003.
Minamino, Tohru – Imada, K. – Namba, K.
(2008): Mechanisms of Type III Protein Export for Bacterial Flagellar
Assembly. Molecular Biosystems. 4, 1105–1115. DOI: 10.1039/B808065H
Muskotál Adél – Seregélyes Cs. – Sebestyén
A. – Vonderviszt F. (2010): Structural Basis for Stabilization of the
Hypervariable D3 Domain of Salmonella Flagellin upon Filament
Formation. Journal of Molecular Biology. 403, 607–615. •
WEBCÍM >
Samatey, Fadel A. – Imada, K. – Nagashima,
S. et al. (2001): Structure of the Bacterial Flagellar Protofilament
and Implications for a Switch for Supercoiling. Nature. 410, 331–337.
DOI:10.1038/35066677
Szabó Veronika – Muskotál A. – Tóth B. et
al. (2011): Construction of a Xylanase a Variant Capable of
Polymerization. PLOS ONE. 6, E25388.
Végh Barbara M. – Gál P. – Dobó J. et al.
(2006): Localization of the Flagellum-Specific Secretion Signal in
Salmonella Flagellin. Biochemical Biophysical Research Communications.
345, 93–98. •
WEBCÍM >
Vonderviszt Ferenc – Kanto, S. – Aizawa,
S. I. et al. (1989): Terminal Regions of Flagellin Are Disordered in
Solution. Journal of Molecular Biology. 209, 127–133. •
WEBCÍM >
Vonderviszt Ferenc – Ishima, R. – Akasaka,
K. et al. (1992): Terminal Disorder: a Common Structural Feature of
the Axial Proteins of Bacterial Flagellum. Journal of Molecular
Biology. 226, 575–579. •
WEBCÍM >
Vonderviszt Ferenc – Imada, K. – Furukawa,
Y. et al. (1998): Mechanism of Self-Association and Filament Capping
by Flagellar HAP2. Journal of Molecular Biology. 284, 1399–1416. •
WEBCÍM >
Vonderviszt Ferenc – Sajó R – Dobó J. –
Závodszky P. (2012): The Use of a Flagellar Export Signal for the
Secretion of Recombinant Proteins in Salmonella. Methods in Molecular
Biology. 824, 134–143. DOI: 10.1007/978-1-61779-433-9_6
Yonekura, Koji – Maki, S. – Morgan, D. G.
et al. (2000): The Bacterial Flagellar Cap as the Rotary Promoter of
Flagellin Self-Assembly. Science. 290, 2148–2152.
DOI: 10.1126/science.290.5499.2148
|