|
használtuk. Ennek lényege az, hogy a Föld felszínén
– azaz a hétköznapi léptékek mellett síkban – egy ismeretlen pont
helyét úgy határozzuk meg, hogy ismerjük három másik, ismert ponttól
való távolságát. Ekkor a három ismert pontból a távolságokkal mint
sugarakkal köröket rajzolva az ismeretlen pont helye a három kör
metszéspontjában lesz. Amennyiben az ismeretlen pontnak csak két másik
ponttól való távolsága ismert, úgy a két kör metszeteként
általánosságban kapható két pont egyikében lesz az ismeretlen pont. A
fehérjék azonban háromdimenziós objektumok, így térben kell
gondolkodnunk, és körök helyett gömböket kell szerkesztenünk.
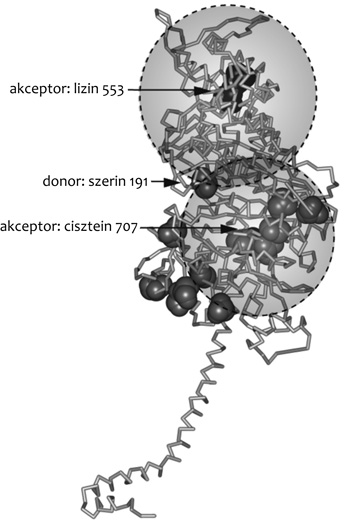
Az ANN tulajdonságait ismerve kiválasztottunk két
olyan további fluorofórt, amelyekkel az ANN energiatranszfer
kapcsolatot hozhat létre. Ezeket korábban már kidolgozott módszerekkel
külön-külön hozzákapcsoltuk a miozinhoz. Mind a két fluorofór esetében
tudtuk tehát azt, hogy hova kötődnek a miozinfejen belül; az egyik egy
cisztein, a másik egy lizin aminosavhoz kapcsolódott. Ezt követően a
FRET módszerének segítségével meghatároztuk, hogy az ismeretlen helyen
lévő ANN milyen távolságban van ezektől a fluorofóroktól, és az ismert
cisztein és lizin pozíciókat középpontnak tekintve, a mért
távolságokkal – mint sugarakkal – megszerkesztettük a két gömböt. A
fentiek szerint, mivel csak két referenciapontot tudtunk alkalmazni,
az ANN helyének pontos kijelölése még egy alulhatározott problémát
jelentett. Ahhoz, hogy ténylegesen és hitelesen megállapítsuk, hova
köt a miozinban az ANN, további ismereteket kellett felhasználnunk.
Ekkor már rendelkezésünkre állt a miozin
szerkezetének atomi pontosságú leírása. A megszerkesztett két gömböt
erre az atomi szerkezetre illesztettük. Szerencsés helyzetben voltunk,
ugyanis kiderült, hogy a két gömb metszeteként előálló kör a
szerkezetben egyetlen szerinmolekulán megy keresztül. A méréseink
alapján tehát ez volt az az aminosav (szerin-181), amelyhez az ANN
kapcsolódott (Szarka et al., 2001). A FRET-módszer alkalmazhatóságát
támasztotta alá, hogy teljesen más eljárást és stratégiát választva
nem sokkal később egy másik kutatócsoport is a miénkkel megegyező
következtetésre jutott az ANN miozinon belüli pozícióját illetően
(Hiratsuka – Katoh, 2003).
Fehérjeszerkezet belső átrendeződésének
vizsgálata: aktin
Az aktin talán „magyar fehérjének” is tekinthető, amennyiben 1942-es
felfedezése és elnevezése a Szent-Györgyi Albert laboratóriumában
dolgozó Straub F. Brunó nevéhez fűződik (Feuer et al., 1948). Az
elmúlt hetven év kutatásai alapján az aktin esszenciálisnak bizonyult
mind az izom-, mind pedig a nem izomsejtek működésében. Biológiai
funkcióját elláthatja monomerként vagy a monomerek specifikus
összekapcsolódását követően filamentális formában is. Szerepe
szerteágazó, bonyolult szabályozási folyamatok által kontrollált.
Ahhoz, hogy ennek a rendkívüli fehérjének a funkcionális sokszínűségét
megérthessük, ismernünk kell, hogy hogyan alkalmazkodik a megváltozott
környezethez, és hogyan befolyásolják a hozzá kötődő kismolekulák vagy
egyéb fehérjék a szerkezetét és a működését.
Az aktinhoz számos más ligandum mellett pozitív
töltésű ionok (kalcium és magnézium) és energiatárolásra képes
molekulák, nukleotidok (ATP) is képesek kötődni. Már a 80-as évek
közepétől külön tudományterület foglalkozott annak leírásával, hogy
ezek a molekulák milyen hatással vannak az aktinra. Vizsgálataink
során mi is tanulmányoztuk a kérdéskört.
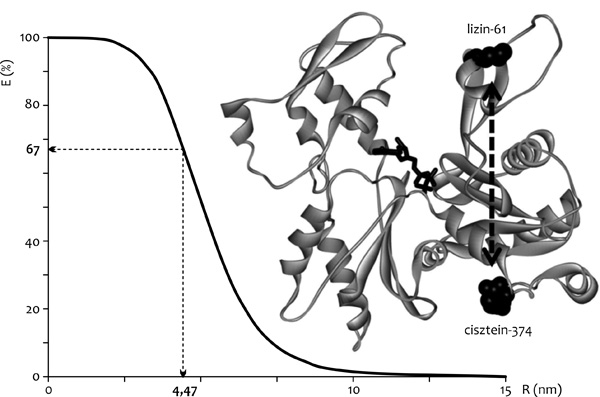
A méréseinket egy speciális, a FRET alkalmazására
épülő módszerre alapoztuk, amelynek alapjait a magyar biofizika egyik
fellegvárában, Debrecenben tették le. Damjanovich Sándor
kutatócsoportjában Somogyi Béla volt az, aki először felvetette annak
a lehetőségét, hogy a FRET alkalmazásával jellemezzük a fehérjék
szerkezeti dinamikájának megváltozását. A módszer használata során a
FRET hatékonyságát több hőmérsékleten is meg kell mérni. A kapott
hőmérsékletfüggő adatok megfelelő kiértékelésével képet kaphatunk
arról, hogy a vizsgált fehérje szerkezeti dinamikai tulajdonságaiban
milyen módosulások álltak be különböző külső körülmények hatására
(Somogyi et al., 1984).
Korábbi tanulmányokból ismert volt, hogy az aktin
funkcionalitása megváltozik különböző kationok vagy nukleotidok
megkötése vagy az oldat savasságának (pH) megváltozása esetén. Mi arra
kerestük a választ, hogy ezek a funkcióbeli különbségek milyen
kapcsolatban vannak az aktin szerkezeti dinamikájának változásával.
Kutatásaink során a fent ismertetett hőmérsékletfüggő FRET-méréseket
alkalmaztuk. A vizsgálatok érdekében az aktint fluoreszcens donorral
(cisztein-374) és akceptorral (lizin-61) jelöltük. A mérési eredmények
ismeretében megállapítottuk, hogy mind az aktin monomer, mind pedig az
aktin filamentum szerkezete merevebb abban az esetben, ha kalcium
helyett magnéziumot köt (Nyitrai et al., 1998; Nyitrai et al., 1999).
Azt is megfigyeltük, hogy az ATP hidrolízisét követően az ADP-t kötő
aktin szerkezete fellazul (Nyitrai et al., 2000), és savasabb
környezetben, azaz alacsonyabb pH-értékek mellett, ez a
fehérjeszerkezet merevebbé válik (Hild et al., 2002).
A FRET-módszer egy speciális alkalmazásával tehát
olyan, eddig nem ismert finom szerkezeti módosulásokat tudtunk leírni,
amelyek az aktin biológiai funkciójának betöltése során fontos
szerepet játszanak.
Ezeknek a kísérleteknek a sikerein felbuzdulva azt
kezdtük el vizsgálni, hogy az aktin szabályozásában részt vevő
fehérjék vajon befolyásolják-e az aktin szerkezetét. A 2000-es évek
elején az aktinkötő fehérjéknek egy speciális csoportja került a
kutatások homlokterébe. Ezek a fehérjék meghatározó szerepet töltenek
be a sejten belüli aktinhálózatok gyors kialakításában. A
filamentumképző fehérjék közül mi a kutatásainkat a forminokra
összpontosítottuk. A hőmérsékletfüggő FRET alkalmazásával kimutattuk,
hogy a forminok által létrehozott aktin filamentumok szerkezete
lazább, mint azoké, amelyek spontán, forminok nélkül alakultak ki
(Bugyi et al., 2006; Papp et al., 2006). A továbbiakban azonosítottunk
két olyan aktinkötő fehérjét, a tropomiozint és a miozint, amelyek a
forminok által fellazított aktin filamentumokat képesek stabilizálni
(Újfalusi et al., 2009).
A vizsgálataink eredményei felvetettek egy izgalmas
kérdést: mi a biológiai szerepe annak a változásnak, amit a forminok
az aktin filamentumok szerkezetében okoznak?
A kérdés megválaszolása érdekében figyelembe
vettük, hogy a sejteken belüli aktinhálózatok vizsgálata során feltárt
apró mozaikokon keresztül kirajzolódott egy érdekes összefüggés. Az
aktinkötő fehérjék egy csoportja azokhoz az aktin filamentumokhoz
kötődik inkább, amelyeket forminok segítségével alakított ki a sejt,
ugyanakkor egy másik csoport nem szívesen kapcsolódik a forminok által
létrehozott filamentumokhoz, sokkal inkább megfigyelhető más
filamentumképző fehérjék környezetében. Ez arra enged következtetni,
hogy az akinkötő fehérjék felismerik, hogy milyen filamentumképző
fehérje hozta létre az aktin filamentumot. Hogy miként? Ez ma sem
teljesen világos.
Eredményeink ismeretében erre az érdekes kérdésre
adódik egy érdekes válasz. Azt a méréseink igazolták, hogy a forminok
megváltoztatják az aktin szerkezetét. Bár még ellenőrzésre vár,
feltételezhető talán az is, hogy más filamentumképző fehérjék is
módosulásokat okoznak az aktin szerkezetében, de ezek más értelmű és
jellegű módosulások, mint a forminok esetében. Ha így van, akkor az is
könnyen elképzelhető, hogy ezek a szerkezeti módosulások befolyásolják
azt, hogy egyes aktinkötő fehérjék milyen erősen tudnak kapcsolódni a
filamentumokhoz. Ez a molekuláris mechanizmus megmagyarázná azokat a
sejtekben tett megfigyeléseket, amelyek az egyes aktinkötő fehérjék
differenciált kötésére vonatkoztak.
A modell jelenleg még igazolásra vár, a folyamatban
lévő kísérletek eredményeinek ismeretében lehet majd hitelesen
alátámasztani az érvényességét. Akár így, akár úgy, ez a példa is
megmutatta, hogy a FRET megfelelő alkalmazásával a biológiai
folyamatok értelmezésének és megértésének új kapui nyílhatnak meg.
Zárszó
A Förster-féle energiatranszfer tanulmányozása és alkalmazása hatalmas
utat tett meg a felfedezése óta eltelt hatvanöt év alatt, a
közelmúltban is számos új, a módszerrel kapcsolatos felfedezés és
elmélet született. Huib Bakker és munkatársai például nehézvízben
karakterizálták az O=D csoportok között végbemenő Förster-féle
energiatranszfert (Piatkowski et al., 2009). Igor Pugliesi és
munkatársai pedig energiatranszfert figyeltek meg egymásra merőleges
tranzíciós dipólmomentummal rendelkező donor–akceptor rendszerekben. A
megfigyelés értelmezéséhez az eredeti Förster-féle képlet módosítására
volt szükség: a szerzők szerint az általuk ismertetett rendszerben az
energiatranszfer hatásfoka a donor–akceptor pár távolságának harmadik
hatványával skálázódik(Pugliesi et al., 2012).
Mindezek fényében optimisták lehetünk a FRET
tekintetében: valószínűleg további innovatív kísérletekre és látványos
eredményekre számíthatunk még hosszú ideig.
Kulcsszavak: fehérjeszerkezet és -funkció, fluoreszcencia
spektroszkópia, Theodor Förster, FRET, aktin, miozin, formin
IRODALOM
Bugyi Beáta – Papp G. et al. (2006): Formins Regulate Actin Filament
Flexibility through Long Range Allosteric Interactions. The Journal of
Biological Chemistry. 281, 16, 10727–10736. •
WEBCÍM >
Feuer György – Molnár F. et al. (1948):
Studies on the Composition and Polymerization of Actin. Acta
Physiologica Hungarica. 1, 4–5,150–163. •
WEBCÍM >
Förster, Theodor (1946): Energiewanderung
und Fluoreszenz. Naturwissenschaften. 6, 166–175. DOI:
10.1007/BF00585226
Förster, Theodor (1948):
Zwischenmolekulare Energiewanderung und Fluoreszenz. Annalen der
Physik. 437, 1–2, 55–75. DOI:10.1002/andp.19484370105
Hild Gábor – Nyitrai M. et al. (2002):
Intermonomer Flexibility of Ca- and Mg-Actin Filaments at Different Ph
Values. European Journal of Biochemistry. 269, 3, 842–849.DOI:
10.1046/j.0014-2956.2001. 02716.x •
WEBCÍM >
Hiratsuka, T. (1989): Nucleotide-Induced
Specific Fluorescent Labeling of the 23-Kda Nh2-Terminal Tryptic
Peptide of Myosin Atpase By the Serine-Reactive Reagent
9-Anthroylnitrile. The Journal of Biological Chemistry.
264(30,18188–18194.
Hiratsuka, Toshiaki – Katoh, Tsuyoshi
(2003): Chemical Identification of Serine 181 at the ATP-Binding Site
of Myosin as a Residue Esterified Selectively by the Fluorescent
Reagent 9-Anthroylnitrile. The Journal of Biological Chemistry. 278,
34, 31891–31894. doi:10.1074/jbc.M303212200 •
WEBCÍM >
Lakowicz, Joseph R. (2006): Principles of
Fluorescence Spectroscopy. Springer, New York •
WEBCÍM >
Nyitrai Miklós – Hild G. et al. (1998):
Effect of Ca2+-Mg2+ Exchange on the Flexibility and/or Conformation of
the Small Domain in Monomeric Actin. Biophysical Journal. 74, 5,
2474–2481. •
WEBCÍM >
Nyitrai Miklós – Hild G. et al. (1999):
The Flexibility of Actin Filaments as Revealed by Fluorescence
Resonance Energy Transfer. The Influence of Divalent Cations. The
Journal of Biological Chemistry. 274, 19, 12996–13001.
doi:10.1074/jbc.274.19.12996 •
WEBCÍM >
Nyitrai Miklós – Hild G. et al. (2000):
Conformational and Dynamic Differences between Actin Filaments
Polymerized from ATP- or ADP-Actin Monomers. The Journal of Biological
Chemistry. 275, 52, 41143–41149. doi:10.1074/jbc.M004146200 •
WEBCÍM >
Papp Gábor – Bugyi B. et al. (2006):
Conformational Changes in Actin Filaments Induced by Formin Binding to
the Barbed End. Biophysical Journal. 91, 7, 2564–2572.
doi: 10.1529/biophysj.106.087775 •
WEBCÍM >
Piatkowski, Lukasz – Eisenthal, K. B. et
al. (2009): Ultrafast Intermolecular Energy Transfer in Heavy Water.
Physical Chemistry Chemical Physics – PCCP. 11, 40, 9033–9038.
DOI: 10.1039/B908975F
Pugliesi, Igor – Langhals, Heinz et al.
(2012): New Perspectives on Ultrafast Förster Resonant Energy
Transfer. XVIIIth International Conference on Ultrafast Phenomena,
Lausanne
Somogyi Béla – Matkó J. et al. (1984):
Förster-type Energy Transfer as a Probe for Changes in Local
Fluctuations of the Protein Matrix. Biochemistry. 23, 15, 3403–3411.
Szarka Krisztina – Bodis E. et al. (2001):
9-Anthroylnitrile Binding to Serine-181 in Myosin Subfragment 1 as
Revealed by Fret Spectroscopy and Molecular Modeling. Biochemistry.
40, 49, 14806–14811. DOI: 10.1021/bi011097k
Ujfalusi Zoltán – Vig A. et al. (2009):
Effect of Tropomyosin on Formin-Bound Actin Filaments. Biophysical
Journal. 96, 1, 162–168. •
WEBCÍM >
Valeur, Bernard (2002): Molecular
Fluorescence : Principles and Applications. Wiley-Vch, Weinheim–New
York •
WEBCÍM >
|