|
|
Az emlősök agykérge tárolja az életük során
összegyűjtött információkat, és mind az állatok, mind az emberek az
elraktározott információ segítségével interpretálják a folyamatosan
érkező kül- és belvilági szenzoros jeleket, miközben agykérgük döntést
hoz a megfelelő motoros programok végrehajtásáról, vagy képzeteket hoz
létre. Az emberben olyan tudati folyamatok is az agykéreg
részvételével mennek végbe, melyeket állatokban többnyire még nem
tudunk mérni: például empátia, szeretet, gyűlölet, szomorúság és hit.
Azt sejtjük, hogy valamilyen módon minden kérgi és tudati folyamat
magyarázata a kérget felépítő idegsejtek jelforgalmának időbeli
összehangolása. A konkrét mechanizmusok meghatározása azonban, vagyis,
hogy az agykéreg időgépezete, a kronocirkuit (krono, idő; cirkuit,
hálózat) az aktuális feladatokat hogyan teljesíti, még mindig óriási
kihívás. Az viszont egyértelmű, hogy a biológiai magyarázathoz
szükséges a kérget felépítő hálózat tér- és időbeli meghatározása.
Milyen mechanizmusok koordinálják időben a kérgi információtároló
idegsejtek aktivitását?
Szentágothai János Michael Arbibbal írt
monográfiájában (Szentágothai – Arbib, 1974) összefoglalta a központi
idegrendszer moduláris szerkezetéről alkotott elképzeléseit.
Szerzőtársával egyebek között az agykéreg működését is szerkezeti és
modellalapon próbálták megjósolni. E mű megelőzi a későbbi és ma is
divatos biológiai alapú agymodellezési próbálkozásokat, de megfelelő
adatok hiányában csak koncepciós vázlatnak tekinthető. Szentágothai
azonban, mint sok más összefoglaló munkájába, ebbe
is belefoglalt máshol nem közölt, eredeti megfigyeléseket. Az agykéreg
neurontípusainak összefoglalásakor leírt, és sematikus rajzzal
illusztrált egy, a macska agykérgében megfigyelt, az irodalomban addig
ismeretlen helyi interneurontípust, amelyet 1975-ben fénymikroszkópos
fotón és részletes rajzon is illusztrált (1.
ábra). A felfedezett sejt axonjának jellegzetes idegvégződései
Szentágothait a csilláron lévő gyertyákra emlékeztették, ezért
kandelábersejtnek nevezte el. Ebben az áttekintésben a
kandelábersejtről szerzett tudásunkon vagy inkább annak
korlátozottságán keresztül azt mutatom be, hogy miért van még mindig
oly kevés ismeretünk agykérgünk, tudatunk és minden abból eredő
kulturális és filozófiai folyamat biológiai magyarázatáról. Mi lehet
az oka annak, hogy felfedezése után harmincnyolc évvel még azt sem
tudjuk a kandelábersejtről, hogy miért kizárólag az agykéregben van rá
szükség, vagy, hogy mi is a szerepe az időhálózatban?
Mint a tudományban oly sokszor, Szentágothai
kandelábersejtjének felfedezése sem elszigetelten történt; tőle
függetlenül, riválisa Edward Jones szintén Golgi impregnációs módszert
alkalmazva majom agykéregben megfigyelt egy hasonló sejtet, melyet
4-es típusú sejtnek nevezett el (Jones, 1975). Visszatekintve
nyilvánvaló, hogy ugyanarról a sejttípusról van szó. Jones is,
Szentágothai is látni vélte, hogy a sejt hatásának kifejtésére az
axonvégződések a piramissejtek apikális dendritjeit célozzák. Jones a
sejt funkcióját illetően nem bocsátkozott feltételezésekbe,
Szentágothai viszont igen: a kandelábersejt gyertyái, azaz gamma-amino
vajsav (GABA) neurotranszmittert felszabadító axonális idegvégződései
többszörös szinapszist létesítenek az agykéreg piramissejtjeinek
apikális dendritjein, és arra gátló hatást fejtenek ki (1. ábra).
Ezt az állítást Szentágothai az axon alakja alapján a tőle megszokott
képzelőerővel jósolta meg, és mivel a nemzetközi porondon rendkívüli
szakmai tekintély volt, állítását senki nem kérdőjelezte meg. A név,
valószínűleg romantikus hangzása miatt, fennmaradt, és ma már senki
nem emlékszik Jones 4-es típusú sejtjére.
A kandelábersejt csak egyike a nagyagykéreg
sejttípusainak, de nem tudjuk, hogy az agyban valójában hányféle
sejttípus van. Ezt azért sem tudhatjuk, mert még elfogadott
kritériumrendszer sincs arra, hogy az agyban egy sejtet mikor
tekintsünk önálló sejttípusnak. Ez az oka annak, hogy az agykéregre
vonatkozó különböző elképzelések igen széles skálán mozognak, húsz és
több ezer közé teszik a különböző sejtféleségek számát. Hát nem
megdöbbentő, hogy minden tudatos pillanatunk agykérgünk terméke, és mi
még azt sem tudjuk, hogy hányféle sejttípus működik
benne?
A továbbhaladás érdekében jelenlegi
munkahipotézisem, hogy az egy sejttípushoz tartozó egyedi sejtek a
biológiai variabilitás és a neuronális plaszticitás határain belül az
agy egy adott állapotában hasonló bemeneti jelkombinációkat hasonló
kimeneti jelkombinációkká transzformálnak. Itt csak a
kandelábersejtnél maradok, de a továbbiak megértéséhez rövid vázlatot
adok bizonyos, az agykéreg szerkezetével kapcsolatos elfogadott
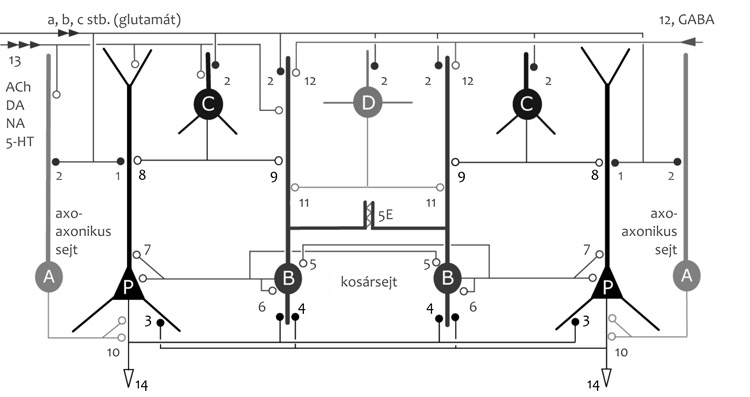
ismeretekről (2. ábra).
Az agykérget többnyire serkentő hatású glutaminsav
(glutamátion) neurotranszmittert felszabadító piramissejtek (kb. 80%)
és legtöbbször gátló hatású GABA-t felszabadító interneuronok (kb.
20%) alkotják, s ezeket legalább öt típusba sorolható gliasejtek
támogatják. A piramissejtek és változataik, mint például a tüskés
csillagsejtek, rétegekbe szerveződnek; az azonos típusú sejtek
többnyire egy rétegben helyezkednek el. Az agykéreg legtöbb területén
hagyományosan hat réteget különítünk el, de ez a kérgi terület
specializációja szerint három és tizenhárom között változhat. A
piramissejtek dendritjei sűrűn tüskések. A többi szinapszistól való
térbeli elkülönítésre a tüskék egy-egy glutamáttal működő szinapszist
alkotnak, melyek hatásfoka és mérete a használat gyakorisága szerint
időben változik. Az információkat, mind rövid, mind hosszú távon (akár
életre szólóan) ezen dendrittüske szinapszisok hatásfokeloszlása
tárolja, de a tüskék meg is szűnhetnek, és újak is képződnek. Azt,
hogy a piramissejtek melyik információtároló vagy -processzáló
idegsejtegyüttes részei, és milyen időbeli aktivitásmintázatokban
vesznek részt, az interneuronok szabályozzák. Az agy állapotának (pl.
alvás, ébrenlét) és a tevékenységnek (például: tanulás, emlékezés) a
függvényében mind a piramissejtek, mind az interneuronok számos kéreg
alatti agyterületekről érkező idegi befolyás alatt állnak. A kéreg
területei között és a kéreg alatti központok sejtjei felé, beleértve
az agytörzsben és a gerincvelőben az izmokat mozgató motoros
neuronokat, az információkat a piramissejtek axonján keletkező és
lefutó akciós potenciálok intenzitása (frekvencia = akciós potenciál
szám/idő) és időbeli mintázata (például: ritmikus működés, csoportos
magas frekvenciájú tüzelés) továbbítja.
Ma már tudjuk, hogy az agykéreg egyik GABA-t
felszabadító interneuronja a kandelábersejt (Freund et al., 1983;
Somogyi et al., 1985; Buhl et al, 1994). Szentágothai állításának
alapja az volt a szerepéről, hogy egy teljesen más agyterületen, a
kisagykéregben a gátló funkciójú kosársejtek szinapszisai hasonló
szerkezeti vonásokat mutattak, mint a Szentágothai által a
piramissejtek apikális dendritjein leírt szinapszisok, melyeket
szimmetrikus szinapszisnak neveztek (összefoglalva: Eccles et al.,
1967). A nagyagykéreg kosársejtjeinek szinapszisai szintén
szimmetrikus szinapszist létesítenek a piramissejt testjein, ezért
ezeket is gátló interneuronnak tartották. A kandelábersejt leírásakor
már ismert volt, hogy mind a kisagyban, mind a nagyagykéregben a
leghatásosabb gátló neurotranszmitter a GABA, ezért a kandelábersejtet
Szentágothai GABA-t felszabadító sejtnek gondolta. A GABA-t
szintetizáló enzimet csak később, 1978-ban mutatta ki Charles E. Ribak
kérgi interneuronokban, de az ismeretlen volt, hogy minden
interneuronban van-e, hogy vajon a kandelábersejt is szintetizál-e
GABA-t, és ha igen, akkor mi a hatása a posztszinaptikus sejtre. Mivel
a piramissejtek radiális irányban futó apikális dendritjén sok
úgynevezett szimmetrikus szinapszis található, és mivel a
kandelábersejt végződései is radiális irányúak, bizonyíték hiányában
is logikusnak látszott, hogy e szinapszisokat a kandelábersejtek
adják.
A 70-es évek elejéig szinte minden agyi szinaptikus
kapcsolatot ilyen indirekt módon jósoltak, mert a Golgi-impregnált (a
kémiailag rögzített sejt ezüstkromát kristállyal való kitöltése
pontosan a plazma membrán által határolt térben), és fénymikroszkópban
azonosított sejtek szinaptikus kapcsolatait ugyanannak a sejtnek az
elektronmikroszkóppal való vizsgálatával rendkívül nehéz volt direkt
igazolni. Ha valaki azt gondolná, hogy a hetvenes években elmaradott
volt a világ, az téved. Ma a genetikailag kódolt fehérjékkel vagy
intracelluláris feltöltéssel láthatóvá tett egyes sejtek közötti
szinaptikus kapcsolatokat legtöbben ugyanilyen indirekt módon, a
fénymikroszkópban látható nyúlványok közeli elhelyezkedése alapján
jósolják meg (lásd kivételek: Biró et al., 2005; Molnár et al., 2008).
Tehát az alkalmazott csúcstechnológiák ellenére még mindig csak
„jóslásokról”, azaz hipotézisekről van szó.
Pályakezdőként elolvasva Szentágothai és Arbib két
évvel korábban megjelent művét, életem céljává az agykéreg kutatását
választottam, s hamar felismertem, hogy a fény- és
elektronmikroszkópos adatok indirekt összehasonlítása rengeteg
bizonytalansághoz vezet. Ezért korábbi próbálkozásokat követve, a SOTE
I. számú Anatómiai Intézetében MTA-segédmunkatársként kifejlesztettem
a fénymikroszkópban azonosított Golgi-impregnált idegsejtek direkt
elektronmikroszkópos szinaptikus vizsgálatát, mely módszer rendkívül
keservesnek, de eredményesnek bizonyult. Az első részletesen vizsgált
Golgi-impregnált sejtem a patkány látókérgében a kandelábersejtre
emlékeztető interneuron volt. Többszöri sikertelen próbálkozás után
egyik késő estén láttam meg a sejt axonja által létesített első
szinapszist, mely a jóslással ellentétben nem a piramissejt apikális
dendritjén, hanem az axonján volt. A váratlan látvány óriási
izgalommal töltött el, és még ma is elfog ugyanez az izgalom, ha arra
az estére gondolok. Miért az izgalom? Mit számít, hogy a piramissejt
axonján vagy dendritjén történik-e a hatás?
Ismert volt, hogy más neuronokhoz hasonlóan, a más
idegsejtektől beérkező sok ezer szinaptikus bemenetet a piramissejtek
is a dendritfájukon integrálják, s azok a sejttest és axon kezdeti
szakaszának membránfeszültségét folyamatosan változtatják. Ha a
membránon keresztül fennálló feszültség elér egy kritikus szintet – az
akcióspotenciál-küszöböt, akkor szinkronizálva kinyílnak a
feszültségfüggő nátriumcsatornák, és tovaterjedő
feszültségváltozás-hullám, akciós potenciál keletkezik, mely a
neuronok közötti jelátvitel alapja. A feszültségfüggő nátriumcsatornák
sűrűsége az axon kezdeti szakaszán (AKSz) a legnagyobb (Lőrincz –
Nusser, 2008), tehát ez a piramissejt kulcsfontosságú jelgeneráló
központja. Mivel a kandelábersejt éppen az AKSz-re koncentrálja az
összes szinapszisát és hatását, nyilvánvaló, hogy a piramissejtek
közötti kommunikáció minden más sejttől eltérő szabályozója. Ez még
ott az első szinapszis azonosítása során átfutott az agyamon. Ez
életem azon ritka pillanatainak egyike volt, amikor valami egy
csapásra megvilágosodott (insight). Az is azonnal eszembe jutott, hogy
mivel egy kandelábersejt több piramissejten ugyanott végződik, azok
jelgenerálását egyszerre befolyásolja, s akármi legyen is az a hatás,
ezzel összekapcsolja sok piramissejt aktivitását. Az
elektronmikroszkópos felvétel negatívjainak előhívása után a fotókat
azonnal lemásoltam pozitív papírképekre, és rohantam Szentágothaihoz.
Diadalmasan mutattam neki, hogy megcáfoltam állítását. A megfigyelés
annyira váratlan volt, hogy eleinte még nem tudhattuk, hogy egy sejt
egy-két szinapszisa alapján mennyire általános az eredmény, de
másnapra több kolléga is megnézte a képeket, és Szentágothai már új
elképzelésekkel érkezett az intézetbe. Meglepetésemre, azt mondta,
hogy ugyan igaz lehet, amit találtam, de ez nem a kandelábersejt,
hanem egy teljesen új sejttípus. Valóban, az általa macska- és
majom-kéregben bemutatott sejtek terminális szakaszai bonyolultabbak
voltak, több végződést tartalmaztak, mint az általam patkányban
vizsgált sejt. Neki is álltam még egy sejtet megvizsgálni, és az is
abszolút specificitást mutatott, csak a piramissejtek AKSz-t
innerválta. Nem volt kétségem, hogy a kandelábersejtről van szó, de
nem tudtam kizárni, hogy azon kívül nem létezik-e még más olyan
sejttípus is, ami az apikális dendritek beidegzésére, innervációjára
specializálódott.
Megírtam a cikket, kézzel kétszer átmásolva adtam
oda Szentágothainak, Somogyi–Szentágothai-társszerzőséggel. Két napon
belül teljesen átírva visszahozta a kéziratot, ő is kézzel írt, de én
már csak egyedüli szerzőként szerepeltem rajta, mert, mint mondta, ő
ehhez az eredményhez nem járult hozzá. A sejtnek az axo-axonikus
(axo-axonal) nevet adta, jelezve a hatás helyének szabályát. A cikk
nagyon megalázó kritikát kapott, a bírálók nem hitték el a
bizonyítékot, de végül 1977-ben megjelent (Somogyi, 1977). Ez máig a
legegyszerűbb közleményem.
Szentágothai a Royal Society kültagja volt, s az
ottani, a Ferrier-díj alkalmából tartott előadásából született cikk
hálózathipotézis rajzában (1/c. ábra) a kandelábersejt és az
axo-axonikus sejt, mint két külön sejttípus szerepelnek a
piramissejtek dendrit-, illetve axonmembránját innerválva
(Szentágothai, 1978), de nincs bizonyíték az apikális dendritet
szelektíven innerváló interneuronra.
Mit tudtunk meg 1977 óta
az axo-axonikus sejtről?
Bármely sejttípus szerepének megértéséhez elengedhetetlen feltétel
meghatározni, hogy a rendszerben honnan kap jeleket, ezek a jelek
mikor érkeznek, hogyan integrálja őket a sejt, az agy állapota szerint
a szinaptikusan befolyásolt sejthez mikor küld ki jelet maga a sejt, s
a jelet milyen molekuláris gépezet viszi át. Az axo-axonikus sejt
jelátadás-helyének felismerése tehát csak az első szerény lépés volt a
megértése felé vezető úton.
Mivel a sejt végződése nagyon jellegzetes és
könnyen felismerhető, az agykéregkutatásban népszerűsége egyre nő,
hetente jelenik meg róla cikk, és már létezik olyan genetikailag
módosított egértörzs, melyben egy medúzából származó zölden
fluoreszkáló fehérjének köszönhetően az axo-axonikus sejtek – és csak
ezek – zölden fluoreszkálnak. A nagy nyüzsgés és érdeklődés ellenére
azonban a tényleges megértés csigalassúsággal halad, s ennek oka
valósággal tükröt tart elénk azzal kapcsolatban, hogy mennyire nem
ismerjük a tudatunkat hordozó agykérgünk működését.
Az axo-axonikus sejt eloszlása az agykéregben
A sejtet leggyakrabban a kéreg II–III. rétegeiben figyelték meg, de a
thalamusból direkt szenzoros bemenetet kapó IV. rétegben és az V–VI.
rétegekben is leírták. Több száz cikk alapján kiderült, hogy ahol
piramissejthez hasonló származású sejtek vannak, például az
amygdalában és a hippokampuszban (például szemcsesejtek), azok minden
emlős fajban, így emberben is kapnak axo-axonikus innervációt is
(Kisvárday et al., 1986). Még nem tudjuk, hogy minden piramissejt
típust innerválnak-e axo-axonikus sejtek; több jel utal arra, hogy
bizonyos típusú piramissejtek működéséhez nem szükséges axo-axonikus
beidegzés. Egy axo-axonikus sejtről egy piramissejtre adott
szinapszisok száma kettő-harminc is lehet (Somogyi et al., 1983a). A
macska látókéregben egy piramissejtre konvergáló axo-axonikus sejtek
száma kb. öt (Freund et al., 1983). Egy AKSz axo-axonikus sejtektől
akár százötven szinapszist is kaphat, ami az agyban a legnagyobb
szinapszissűrűségek közé tartozik, jelezve, hogy nagy hatásfokú
kölcsönhatás része. Egy axo-axonikus sejt 200–1200 piramissejtet is
innerválhat, mindegyiket az AKSz-n (Li et al., 1992).
Az axo-axonikus sejt neurotranszmittere
és a posztszinaptikus receptorok
Az axo-axonikus sejt jelátvivő transzmitterére az első direkt
bizonyítékot Freund Tamás szolgáltatta (Freund et al., 1983), amikor
Golgi-impregnációval azonosított axo-axonikus sejt végződéseiben
elektronmikroszkópos immuncitokémiai módszerrel kimutatta a GABA-t
szintetizáló enzimet, a GAD-ot. További indirekt bizonyítékok
után végül magát a GABA-t Freund Tamással együtt
mutattuk ki Golgi-impregnált axo-axonikus sejtben a
macska-hippokampuszban, 1985-ben (Somogyi et al., 1985). Az
axo-axonikus sejt terminálisaiból felszabaduló GABA piramissejtre
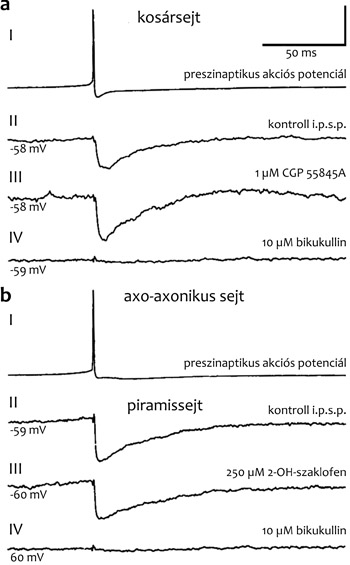
gyakorolt hatását Eberhard Buhl mérte először (Buhl et al., 1994),
felnőtt patkány-hippokampusz túlélő szeletben intracelluláris
elektródával elvezetve mind a preszinaptikus axo-axonikus sejtet, mind
a posztszinaptikus piramissejtet (3. ábra).
Mivel mai tudásunk szerint a hippokampuszban GABA legalább
huszonegyféle interneuronból szabadulhat fel (Somogyi, 2010), fontos
volt, hogy Buhl az üveg kapilláris elektródán keresztül az elvezetett
sejteket megjelölte, és így a sejt formájáról és terminálisainak
szinaptikus helyéről igazolni tudta, hogy axo-axonikus sejt hatását
mérte. Az axo-axonikus sejtben lévő üvegelektródán keresztül a
véletlenül talált egyedi preszinaptikus sejtet tetszés szerint tudta
ingerelni. A kiváltott akciós potenciálokkal a piramissejt testében
(szómájában) gyors hiperpolarizációt figyelt meg, amit az A típusú
GABA receptor antagonista növényi alkaloida, a bikukullin, teljesen
megszüntetett (3. ábra). Ezzel bizonyítottá vált, hogy az
axo-axonikus sejt hatását GABA-A-receptoron keresztül fejti ki, mely
egy több alegységből álló anioncsatorna. Egyébként ez a benzodiazepin
nyugtató és altató gyógyszercsalád (pl. Valium, Librium) hatásának
helye. A sok GABA-A-receptor altípus közül az alfa-2-es alegységet
tartalmazó receptor, melyen a szorongást csökkentő benzodiazepinek is
részben hatnak, különösen nagy mennyiségben fordul elő az axo-axonikus
sejt szinapszisokban (Nusser et al., 1996). Mivel azonban ezek a
szerek még sok helyen fejtik ki hatásukat, nem tudni, hogy az
axo-axonikus sejtnek van-e köze a szorongáscsökkentő effektushoz.
További, a sejt működési mechanizmusára utaló
jellemzők, hogy az axo-axonikus sejt rövid időtartamú akciós
potenciálokat generál, és egyike azoknak a kérgi sejteknek, amelyek
magas akcióspotenciál-frekvenciára képesek; az axo-axonikus sejt ún.
gyorstüzelő sejt; ezt a membránjában lévő feszültségfüggő csatornák
teszik lehetővé. A legtöbb axo-axonikus sejt a teljes sejtben, így az
axonvégződésekben is parvalbumin nevű kalciumkötő fehérjét tartalmaz,
s ez kalciumpufferként hatva lehetővé teszi, hogy az egymás után
érkező akciós potenciálok rövid idő alatt szabadítsák fel a GABA-t,
azaz a transzmitterfelszabadulás pontosan tükrözze a sejt tüzelését.
Ez precízen időzített jelátvitelre utal.
A fordulat – piramissejtek precíz tüzeltetése
axo-axonikus sejttel in vitro
Bár senki nem kételkedett az axo-axonikus sejtek gátló hatásában, arra
mégsem volt magyarázat, hogy miért éppen a piramissejteknek kell olyan
gátló partner, amely csak az axonon tovaterjedő akciós potenciál
kiváltó helyen, az AKSz-n hat. A piramissejttest kosársejtektől
rengeteg GABA-t felszabadító szinapszist kap, és a dendriteket
legalább egy tucat különböző típusú és GABA-t felszabadító interneuron
innerválja (2. ábra). Az, hogy az AKSz külön forrásból kap
GABA-t, arra utal, hogy a felszabadulás időbeli dinamikája eltér a
többi preszinaptikus sejtétől, s ezért a GABA külön sejtből kell
származzon, és a sejt egyes részein eltérő módon fejtheti ki hatását.
Szegeden, Tamás Gábor laboratóriumában túlélő kéregszeletben Szabadics
János egyszerre több kérgi neuron membránpotenciálját figyelte, és
meglepő módon néha azt találta, hogy amikor egy GABA-t felszabadító
sejtben akciós potenciált vált ki, a várt hiperpolarizációs gátló
hatás helyett egyik-másik kérgi sejtben depolarizáló szinaptikus
választ, glutamátreceptor aktiválásból származó serkentő
posztszinaptikus potenicált (EPSP) kapott, mely csak piramissejttől
származhatott. Az évek alatt összegyűlt hasonló, ritka minták
fénymikroszkópos vizsgálata során Tamás Gábor észrevette, hogy a
preszinaptikus sejt minden ilyen különös esetben axo-axonikus sejt
volt. Azaz, az axo-axonikus sejtből felszabaduló GABA valahol a
szeletben piramissejtet aktivált. Tamás Gábor vezetésével Szabadics
János és Molnár Gábor, a preszinaptikus interneuronokat és a
posztszinaptikus piramissejteket anatómiailag is azonosítva több sejt
szimultán elvezetésével részletesen összehasonlították az axo-axonikus
és a szómán végződő kosársejt hatását (Szabadics et al., 2006).
Meglepő módon azt találták, hogy a GABA-A-receptorcsatornán át töltést
hordozó kloridionok egyensúlyi potenciálja az AKSz-ben jóval
pozitívabb, mint a szómában, s ennek következtében néha az
axo-axonikus sejtben kiváltott akciós potenciállal, ami az AKSz
szinapszisaiban GABA-t szabadít fel, bizonyos piramissejteket minden
addiginál precízebben lehet tüzeltetni. Ezt azonban csak úgy lehetett
elérni, ha a sejt belső citoplazmatikus oldatát az elvezető
üvegelektródában lévő mesterséges sóoldatra cserélték le.
A piramissejtek tüzelését azzal magyarázták, hogy a
megnyíló GABA-A-csatornákon a piramissejtből kifolyó klorid
depolarizálja az AKSz-t, és akciós potenciált generál. Ezzel
összhangban, szemben a sejttest plazmamembránjával, az AKSz-n Varga
Csaba elektronmikroszkópos immunohisztokémiai módszerrel alig talált
olyan fehérjét, mely kloridionokat pumpál a a sejtből kifele, azaz az
AKSz-ben magasabb lehet a kloridion-koncentráció, mint a szómában. A
szómán végződő kosársejt szinapszisaiból felszabaduló GABA-val nem
tudtak a piramissejtből akciós potenciált kiváltani, mert a klorid
egyensúlyi potenciálja a tüzelési küszöbhöz képest negatívabb volt, és
az alacsonyabb nátriumcsatorna-sűrűség miatt a tüzelési küszöb értéke
is pozitívabb sejttesten. Részletes kísérleteikből arra a
következtetésre jutottak, hogy az axo-axonikus sejtek bizonyos
körülmények között serkenthetik a piramissejteket, és egy axo-axonikus
sejt által innervált piramissejt-populációban szinkronizált tüzelést
válthatnak ki. Kísérleteiket agyműtét során eltávolított túlélő emberi
agykéregszeletben is sikerrel megismételték (Szabadics et al., 2006).
Ez az első olyan eset, amikor emberi axo-axonikus sejt fiziológiai
hatását sikerült azonosítani. További kísérleteik során Tamás és
munkatársai kimutatták, hogy az emberi agykéregben egyedülálló módon
egyetlen idegsejt ingerlésével szinaptikus események hosszú láncolata
váltható ki, amelyek közvetítésében szintén axo-axonikus sejtek
részvételét valószínűsítették (Molnár et al., 2008). Tehát, az AKSz-t
innerváló axo-axonikus sejt egyetlen akciós potenciállal időhálózati
mintát generált, nem pedig csak gátolt, mint korábban állítottuk. Az
AKSz-n történő GABA által kiváltott depolarizációs hatást és
piramissejt-tüzelést in vitro több csoport is megerősítette, és
hiperpolarizációról is jelentek meg munkák. Szabadics és Tamás szerint
a piramissejtre az axo-axonikus sejt által kifejtett hatás a sejtet
érő egyéb hatásoktól függően dinamikusan változhat, mivel a GABA által
kiváltott membránválasz egyensúlyi potenciálja közel van a tüzelési
küszöbhöz. Elképzelésük szerint az axo-axonikus sejtek a célsejtek
döntő többségén gátló hatásúak, és csak serkentő funkcióban, a
célsejtek egy kis csoportján (néhány százalékán) működhetnek. Így a
hálózatban megmaradhat a serkentés-gátlás egyensúlya, és emelt jel-zaj
viszonyú jelsorozatok képződhetnek. Itt jegyzem meg, hogy a GABA által
kiváltott depolarizációs hatás az AKSz-n nem szükségszerűen serkentő,
lehet gátló, sőt erősen gátló is, mivel a receptorcsatornák
nyitottsága alatt söntölő (vagy „csendes”) gátlást képezve csökkenti
az AKSz bemenő ellenállását, és ez csökkenti a sejttest felől érkező
depolarizáció akcióspotenciál-generáló hatékonyságát. Még fontosabb
lehet, hogy az axo-axonikus sejt által kiváltott viszonylag lassú
depolarizáció az akciós potenciálok kialakulásáért felelős
nátriumcsatornák egy részét inaktiválhatja, csökkentve ezáltal ezen
csatornák nyithatóságát, ami végső soron ahhoz vezet, hogy a
piramissejt
|
|
|
nehezebben képes akciós potenciált generálni, tehát
gátolt. Hasonló mechanizmus működik a gerincvelő hátsó szarvában lévő
primer szenzoros afferensek terminálisain lévő GABA-t felszabadító,
GABA-A-receptort használó axo-axonikus szinapszisokban is. Az
axo-axonikus sejt tehát hiperpolarizációval, depolarizációs sönttel és
nátriumcsatorna-inaktiválással többszörös mechanizmuson keresztül is
gátolhatja a piramissejt működését.
A piramissejtből depolarizációval kiváltott
akcióspotenciál-generálás, mely az axo-axonikus sejt in vitro mért
serkentő hatására utal, vajon összeegyeztethető-e az axo-axonikus sejt
általi gátló hatásra mutató adatokkal? Elképzelhető-e, hogy a sejt
állapotától függően mindkét hatás lehetséges? Egyelőre egyik effektus
sem zárható ki, de nyilvánvaló, hogy a működő agyban a hatás attól
függ, hogy amikor GABA az AKSz-n lévő receptorokra érkezik, mennyi a
piramissejt membránpotenciálja és bemenő ellenállása. Ez in vitro jól
mérhető, de nem tudni, hogy ezek a körülmények mennyire tükrözik az
élettani állapotot, mert a sejten kevés szinapszis működik; a
mesterséges sóoldatban mások az extracelluláris
transzmitter-koncentrációk és a sejtek az in vivo működő rendszerhez
képest keveset tüzelnek spontán. Viszont, abban a pillanatban, amikor
az AKSz GABA-t kap, in vivo szinte lehetetlen a membránpotenciált és a
bemenő ellenállást mérni. Ha az AKSz GABA-A receptorainak szinaptikus
aktiválása az adott pillanatban piramissejtben valóban kiváltana
akciós potenciált, akkor a túlélő szeletben az akciós potenciál
hatására belépő kálciumionok optikai módszerrel történő kimutatásával
ezeket a tüzelő piramissejteket detektálni lehetne. Az eddigi
próbálkozások nem vezettek eredményre, de ezzel a módszerrel egy rövid
időablakban lehetetlen minden piramissejtet követni. Összefoglalva:
arra nincs bizonyíték, hogy az axo-axonikus sejtek in vivo körülmények
között képesek lennének a piramissejt akcióspotenciál-generálásának
valószínűségét növelni (serkentés), de ez nem is zárható ki.
Egyik hipotézisem, hogy a piramissejtek AKSz-én
GABA-szinapszison keresztül in vitro kiváltott akciós potenciálok
(Szabadics et al., 2006; Molnár et al., 2008), melyeket kétség kívül
többen is demonstráltak, olyan AKSz-ből származnak, melyekhez tartozó
piramissejt az agyszelet készítése során megsérült, vagy talán az
axonhoz már nem is tartozik sejttest, mivel az elvágott axonok
lezáródnak, de még órákig működnek. A sejttest és a dendritfa
kapacitásterhelésétől megszabadult túlélő axont az axo-axonikus sejt
szinapszisai a szómán lévő kloridpumpa intracelluláris
kloridkoncentrációt csökkentő hatásának hiányában nagy hatásfokkal
depolarizálhatják. Így amíg a piramissejt axonja él, a szeletben
hatékony serkentő kapcsolatokat tarthat fenn.
Az axo-axonikus sejt működése in vivo –
ritmikus kérgi időminták
Az összeköttetésbeli, biofizikai és farmakológiai eredmények fontosak
a sejtszintű mechanizmusok tisztázására, de nem elegendőek annak a
kérdésnek a megválaszolására, hogy az agykéregben az axo-axonikus sejt
mire is való. Mindenki sejti, hogy valamilyen sok piramissejtet
koordináló hatása lehet, és erről rengeteg spekuláció született,
amelyek némelyike talán igaz is. Viszont a valós szerep
megközelítésére a teljes működő rendszerben kellene megfigyelni az
axo-axonikus sejteket. Sokáig a kérgi neuronok között csak
véletlenszerűen lehetett axo-axonikus sejtet találni, s mivel az
összes neuron kevesebb mint egy százalékát alkotják, és ráadásul a
kisebb méretű sejtek közé tartoznak, valóságos csoda, hogy az
azonosításhoz szükséges mikroszkópos felismerésre történt sikeres
elektrofiziológiai elvezetés és ezt követő sejtjelölés történt. Teljes
állatban axo-axonikus sejtet először Kevan Martin jelölt Oxfordban
intracelluláris hegyes üvegelektródával macska-látókéregben, és a
sejtet torma-peroxidáz enzim beadásával tette láthatóvá (Freund et
al., 1983). Sajnos a sejt vizuális ingerre adott válaszát nem tudta
meghatározni, mert az axo-axonikus sejt mellett egy piramissejt is
jelölődött, így nem lehetett megállapítani, hogy az elvezetett akciós
potenciálok melyik sejttől származnak. Az első hippokampális in vivo
azonosított axo-axonikus sejtet hasonló balszerencse érte (Li et al.,
1992).
A patkány szomato-szenzoros kéregben végzett
intracelluláris axo-axonikus sejtelvezetések ugyan több paraméterben
más választ mutattak, mint a többi, GABA-t felszabadító interneuron,
de az alkalmazott barbiturát altatás miatt a sejt szerepéről kevés
derült ki (Zhu et al., 2004).
Végül 2003-ban jelent meg az első beszámoló
uretánnal altatott állatban azonosított axo-axonikus sejtek tüzelési
mintázatáról a hippokampuszban (Klausberger et al., 2003).
Viselkedéssel járó agyi aktivitást ugyan az alkalmazott altatás miatt
nem lehetett tesztelni, de mivel a hippokampuszra jellemző agyi
ritmikus aktivitást, mint például a théta frekvenciájú (4–8 Hz) vagy
az éles hullámhoz kapcsolt nagyfrekvenciás mezőpotenciál-oszcillációt
(130–200 Hz) ez az altató nem szünteti meg, ezen oszcillációk időbeli
referenciát adnak az axo-axonikus sejt hálózati működési időpontjairól
a többi sejthez képest. Az éles hullám egy nagy amplitúdójú
extracellulárisan detektált mezőpotenciál, melyet Buzsáki György
fedezett fel a hippokampuszban, és kiderítette, hogy azt a CA3-régió
piramissejtjeinek szinkronizált tüzelése hozza létre mintegy 50–150 ms
időtartam alatt (Buzsáki, 2006). Ezek a ritmikus kérgi aktivitásminták
szinkronizált piramissejt-működést jeleznek, és viselkedésfüggőek. Így
a hippokampuszban a théta-oszcilláció a mozgással járó
információkódolás és memórianyomok előhívását jelzi, míg a főleg alvás
alatti éles hullám az emléknyomok hippokampuszból az agykéregbe
történő beolvasását teszi lehetővé (Buzsáki, 2006). Az eltérő
frekvencia- és időskála ellenére, mindkettő lényege, hogy a
piramissejtek vagy 100–200 milliszekundumonként (théta) vagy 5–7
milliszekundumonként (éles hullám) ritmikusan gátoltak, ami azzal jár,
hogy a gátlás közötti időablakokban nagyobb
valószínűséggel működnek együtt. A ritmikus gátlás tehát az
információkódolás, -rögzítés és -előhívás kulcsa, mert időben
összehangolja az egymással kapcsolatban lévő sejteket, melyek részt
vesznek az emléknyomok agyi reprezentálásában.
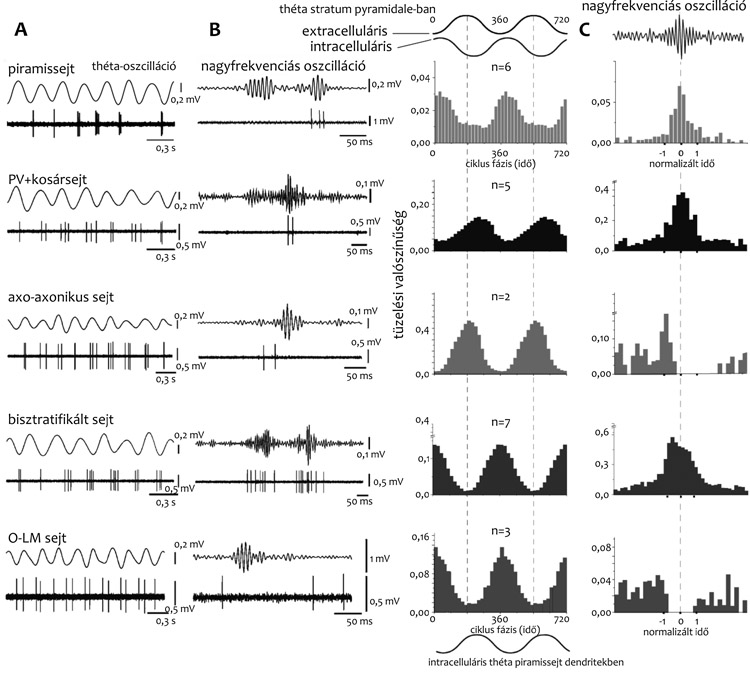
Vajon a piramissejt mely részén történik ritmikus
gátlás? Thomas Klausberger és munkatársai mintegy tizenöt, GABA-t
felszabadító interneuronfajta tüzelésének in vivo elvezetésével
felfedezték, hogy az időhálózatban mindegyiknek saját időbeli
specificitása van (4. ábra, példák). Bár
a legtöbb GABA-t felszabadító sejtfajta ritmikusan működik, itt csak
az axo-axonikus sejt ritmusára hívom fel a figyelmet, mely a
piramissejt-rétegben detektált extracelluláris mezőpotenciál théta
hullám pozitív csúcsán tüzelt (Klausberger et al., 2003). Mind az
altatott, mind a szabadon mozgó állatban ez a ritmus azon fázisa,
amikor a piramissejtek legkevesebbet tüzelnek, és a legkevésbé
ingerelhetők, mert gátoltak. Az axo-axonikus sejt maximális tüzelése
tehát egybeesik a piramissejt tüzelésének maximális gátlásával. Ez
talán nem hangzik meglepőnek, de mint a 4. ábrán látszik, más
GABA-t felszabadító sejtek, mint például a bisztratifikált sejt és az
O-LM-sejt, éppen a piramissejtek: a théta hullám legnegatívabb
periódusa körül tüzelnek maximálisan. Ebben a cikkben most nincs
helyem ennek a látszólagos paradoxonnak a magyarázatára.
A másik ritmus, a nagyfrekvenciájú oszcilláció
alatt az axo-axonikus sejtek gátoltak (4. ábra, Klausberger et
al., 2003). Ez a nagyfrekvenciájú oszcilláció az agykéreg egyik
legpontosabb és legnagyobb méretű, időben összehangolt
piramissejt-kisülése, mely alvás alatt 50–100 ms időtartamban
visszajátssza azt a piramissejt-aktivitássorrendet, amit az állat több
másodperc alatt éberen átélt (Buzsáki, 2006). Ezt úgy is lehet
értelmezni, hogy a piramissejt-együttesek visszajátszanak egy
emléknyomot, de csak a piramissejtek azon kis része vesz részt egy
ilyen kisülési hullámban, amely az adott emléknyomot kódolta (Buzsáki,
1989). Buzsáki György állította fel azt a hipotézist, hogy ez azért
következhet be, mert a piramissejtek gátlása ebben az 50–100 ms-os
periódusban csökken, azaz a piramissejtek ingerelhetőbbé válnak, és
Csicsvári Józseffel talált is olyan nem azonosított interneuronokat,
melyek az éles hullám alatt elhallgattak (Buzsáki, 2006).
Klausberger és munkatársai (2003), köztük Buzsáki
és én, azonosították az első ilyen interneuront, mely axo-axonikus
sejt volt. Tehát ez a GABA-t felszabadító sejt visszavonja hatását a
piramissejt AKSz-ról, pont onnan, ahol a piramissejtek akciós
potenciálja keletkezik, s így az axo-axonikus sejt gátlása hozzájárul
a piramissejt együttes kisülési feltételéhez. Ezek a mérések a
hippokampusz CA1-régiójából származnak, viszont az itt tüzelő
piramissejteket az éles hullámot generáló CA3-as régió pozitív
visszacsatolásra vezető, szinaptikusan sűrűn összekötött
piramissejtjei serkentik. A megértés kulcsa tehát annak ismerete, hogy
a CA3-as régió axo-axonikus sejtjei is gátoltak-e az éles hullám
alatt, de erre nincs adat. Lasztóczi Bálint, John Tukker és Thomas
Klausberger Oxfordban végzett, még nem közölt mérései azt mutatják,
hogy ezen összehangolt populációs tüzelés alatt a CA3-as régió éles
hullámot generáló piramissejtjeit innerváló axo-axonikus sejtek is
valóban gátoltak. Ezek alapján elképzelhető, hogy lassú hullámú alvás
alatt az axo-axonikus sejt gátlása az éles hullám alatti
memóriamegerősítés feltétele.
Összefoglalva: az axo-axonikus sejtek
hippokam-pális időhálózati működése azt mutatja, hogy az axo-axonikus
sejtek akkor tüzelnek a legjobban, amikor a piramissejtek a legkevésbé
(théta csúcs), és akkor gátoltak, amikor a piramissejtek a legjobban
tüzelnek (éles hullám/nagy frekvenciájú oszcilláció). Ez a a
piramissejt és az axo-axonikus sejt tüzelése között fennálló kettős
fordított korreláció indirekt bizonyítéknak tekinthető az axo-axonikus
sejt gátló hatására. A théta aktivitás alatti ritmikus
piramissejt-gátlással az axo-axonikus sejt a piramissejt együttesek
időbeli szekvenciasorait teszi lehetővé, míg a gátlás visszavonása az
éles hullámhoz kapcsolt nagyfrekvenciájú oszcilláció alatti
visszajátszás alatt az emléknyomok rögzítésére vezethet.
Általánosabban, az axo-axonikus sejt a piramissejt-tüzelés egyedülálló
szabályozása révén az agykérgi aktivitás időmintáinak kulcsfontosságú
szabályozója.
Tudásunk korlátai – mely bemenő jelek aktiválják vagy gátolják az
axo-axonikus sejtet
Ha a fenti spekuláció megfelel a valóságnak, akkor alapvető kérdés,
hogy mitől tüzel az axo-axonikus sejt a fenti minták szerint, azaz
milyen serkentő vagy gátló innerváció időbeli dinamikája befolyásolja.
Erről tudunk a legkevesebbet, és ez nemcsak erre a sejtre, hanem az
agykéreg szinte valamennyi neuronjára vonatkozik. Nem ismerjük, hogy
mely gátló és serkentő bemenetek vezetnek egy adott kérgi neuron
membránponteciáljának akcióspotenciál-küszöb elérésére. De még a
listáink is nagyon hézagosak a fizikailag azonosított bemenetekről,
nem is beszélve azok fiziológiai dinamikájáról. Az axo-axonikus sejt
esetén helyi piramissejtekkel történő páros elvezetéssel és
sejtjelöléssel bizonyították, hogy utóbbiaktól serkentő szinaptikus,
sőt reciprok kapcsolatban állnak (Ganter et al., 2004; Molnár et al.,
2008). Hogy ez mekkora részét képezi a serkentő bemenetüknek, nem
ismert. A hippokampusz glutamáttal működő innervációjának réteges
eloszlásából és az axo-axonikus sejtek dendritjeinek ezekkel való
átfedéséből közvetve ugyan, de biztosra vehető, hogy ott mind távoli
neokortikális, mind intrahippokampális piramissejtekből jövő serkentő
bemenet nagyobb súllyal szerepel, mint az adott axo-axonikus sejt
által innervált piramissejtektől jövő reciprok bemenet (Somogyi et
al., 1985; Klausberger et al., 2003). Valószínű, de még tesztelésre
vár, hogy az axo-axonikus sejt dendritfáját a terében végződő összes
glutamátpályák szinaptikusan innerválják-e. Ebből viszont sem azok
relatív súlyát, sem időbeli hozzájárulását a sejt aktiválásához nem
tudjuk. Még siralmasabb a kép az axo-axonikus sejteken végződő gátló,
vagy GABA-t használó szinaptikus innervációról. Tudomásom szerint erre
csak Tamás Gábor munkacsoportjának vannak eddig még nem közölt
mérései, közleményt pedig nem ismerek. Az agykérget innerváló többi
kéreg alatti agyterületnek, mint például a thalamusnak vagy a monoamin
pályáknak az axo-axonikus sejthez való viszonya szintén ismeretlen.
Tehát az axo-axonikus sejt agykéregben játszott
valós szerepét nem tudjuk megállapítani, amíg a működését szabályozó
szinpatikus bemenetek ismertté nem válnak. A többi agykérgi sejttípus
sem áll sokkal jobban. Pedig a szinaptikus bemenetek megállapítására
vannak módszerek. Hogyan lehetséges, hogy bár az axo-axonikus sejt
fontos lehet a tudatunk alapját képező piramissejt tüzelési időmintái
felállításában, felfedezése után harmincnyolc évvel még mindig ilyen
kevés az ismeret? Az ismeretek hiánya jól tükrözi az emberi
pszichikumot, ami szintén az agykéreg terméke. Össztársadalmi szinten
fontosabbnak tartjuk pusztító fegyvereinket még pusztítóbbakra
lecserélni, óriási összegekkel államilag támogatni környezetkárosító,
ám politikailag látványos és egyeseknek előnyös gazdasági
tevékenységeket, mint megtudni, hogyan is működik az a kérgi hálózat,
amely ezeket a tevékenységeket lehetővé teszi. Kérgünk, tudatunk
egyszerűen van, s társadalmunk csak akkor szán rá figyelmet, illetve
némi aprópénzt, amikor valamiért nem jól működik.
Lehetőségek és remények azonosított típusú idegsejtek szerepének
magyarázatára
és fennmaradásunkra
Az elmúlt két évben az agykéreg és a teljes agy időhálózatának
meghatározására minden korábbi elképzelést felülmúló lehetőségek
nyíltak.
1. A genetikailag módosított, fluoreszkáló
veszettségvírus transz-szinaptikus transzportjának zseniális
kihasználásával egy intracellulárisan in vivo élettanilag jellemzett
és elektrofiziológiailag elvezetett posztszinaptikus sejtből, a sejtet
innerváló preszinaptikus neuron megjelölésével (Rancz et al., 2011)
elvileg lehetővé vált a teljes agyban az egy axo-axonikus sejtet
innerváló összes sejt anatómiai feltérképezése.
2. Az axo-axonikus vagy más kiválasztott sejttípus
zöld fluoreszkáló fehérjével genetikailag szelektíven való megjelölése
lehetővé teszi, hogy mikroszkóp alatt fejrögzített, nem altatott és
viselkedési tesztet végző egér agykérgében a kiválasztott sejt
működését kövessék a kutatók, miközben mikroszkóp alatt optikai módon
kálcium leképezéssel több száz vagy ezer egyedi kérgi sejt tüzelését
is detektálhatják. A viselkedés alatt a kiválasztott, például
axo-axonikus sejt aktivitását akár intracellulárisan is el lehet
vezetni (Gentet et al., 2012; Z. Joshua Huang, személyes közlés
axo-axonikus sejtre).
3. Megvalósult szabadon mozgó patkányok egyes
hippokampális interneuronjainak viselkedés- és hálózatállapot-függő
tüzelésének követése, majd a neuron megjelölése az állat fején
hordozott miniatűr üvegelektródát hajtó motor segítségével (Lapray et
al., 2012). Tudomásom szerint axo-axonikus sejtet még senki nem jelölt
így, de ez csak idő kérdése.
E technikák persze csak további, de most már az
egész rendszerre jellemző adathalmazt produkálhatnak. A tényleges
megértésbeli áttöréshez szükség van az adatkezelő elveket és a
biológiai adatokat pontosan használó új matematikai modellek
fejlesztésére, mely, ha meglehetősen sok zsákutcával és valóságtól
elszakadt hírveréssel is, de szintén gyorsan halad előre. Jelenleg a
legfontosabb feladat a valós hálózatok által végzett
információfeldolgozó lépések (computation) formalizálása.
És miben reménykedem? Remélem, még megélem, hogy
felfedezésének 50. születésnapján, ami Szentágothai János születésének
112. évfordulója lesz, az axo-axonikus sejtről összeáll egy kép, és
kiderül, miért nem működik nélküle az agykéreg. Ehhez fenn kellene
maradnia a természet ismeretén, a tudományos világszemléleten alapuló
racionális és technológiai társadalomnak, melynek fennmaradásában
Szentágothai nem volt biztos. Ő elsősorban az atomkatasztrófától
tartott.
Eltávozása óta a helyzet nem javult ezen a téren,
mert bár atomháborútól manapság nem kell félnünk, több oldalról is
megtudtuk, hogy agykérgünk rendkívüli képességeinek felhasználásával
ez a társadalom mely más utakon pusztíthatja el magát. A tudományos
bizonyítékon alapuló felismerések kapcsán, ilyen például az ember
által okozott klímaváltozás, megindultak a társadalmi viták is, de
elsősorban kérgi tudati mechanizmusaink korlátai miatt, a konkrét
lépések ugyanolyan csigalassúsággal születnek, mint amilyen lassan az
agykéreg kutatása halad.
Szentágothai formafelismerő zsenialitása
felkapcsolta a kandelábert az agykéregben. Ő felállított egy
munkahipotézist, mely azóta módosult, és ma már több százan dolgozunk
tovább rajta. Közülünk többen nem is ismerik az ő hozzájárulását. Ez a
tudományos haladás törvényszerűsége, de mint mondtam egy
Szentágothairól adott nyilatkozatomban „…szellemi csillagpora itt él
tovább kultúránkban, tudásunkban, az egyetemes emberi tudományos
haladás halhatatlan hagyatékaként.” (Kittel, 2012)
A kandeláber fénye egyre erősödik, mert, ahogy az
embert az állatok viselkedésének tanulmányozásán keresztül megérteni
akaró Nobel-díjas Konrad Lorenz írja: „A tudományos igazság a legjobb
munkahipotézis, mely megnyitja az utat a következő még jobb felé.”
Köszönöm Dr. Szabadics Jánosnak, Dr. Tamás Gábornak és Dr. Somogyi
Józsefnek a cikkel kapcsolatos javaslatait a kézirat egy korábbi
változatához, és Gimes Júlia szerkesztői segítségét és nyelvi
javításait a kéziratban.
Kulcsszavak: agykéreg, neuronális aktivitás, gátlás, szinapszis,
akciós potenciál, elektrofiziológia
IRODALOM
Biró Ágota A. – Holderith N. B. – Nusser
Z. (2005): Quantal Size Is Independent of the Release Probability at
Hippocampal Excitatory Synapses. The Journal of Neuroscience. 25,
223–232. • doi:10.1523/JNEUROSCI.3688-04.2005 •
WEBCÍM >
Buhl, Eberhard H. – Halasy K. – Somogyi P.
(1994): Diverse Sources of Hippocampal Unitary Inhibitory Postsynaptic
Potentials and the Number of Synaptic Release Sites. Nature. 368,
823-828. doi:10.1038/ 368823a0
Buzsáki György (1989): Two-Stage Model of
Memory Trace Formation: A Role for ‚Noisy’ Brain States. Neuroscience.
31, 551–570. •
WEBCÍM >
Buzsáki György (2006): Rhythms of the
Brain. Oxford University Press, New York •
WEBCÍM >
Eccles, John C. – Ito, M. – Szentágothai J. (1967). The Cerebellum as
a Neuronal Machine. Springer, Berlin
Freund Tamás F. – Buzsáki György (1996):
Interneurons of the Hippocampus. Hippocampus. 6, 347–470. •
DOI: 10.1002/(SICI)1098-1063(1996)6:4<347::AID-HIPO1>3.0.CO;2-I
Freund Tamás F. – Martin, K. A. C. –
Smith, A. D. – Somogyi P. (1983): Glutamate
Decarboxylase-Immunoreactive Terminals of Golgi-impregnated Axo-axonic
Cells and of Presumed Basket Cells in Synaptic Contact with Pyramidal
Neurons of the Cat’s Visual Cortex. The Journal of
Comparative Neurology. 221, 263–278. • DOI: 10.1002/cne.902210303
Ganter, Paul – Szücs P. – Paulsen, O. –
Somogyi P. (2004): Properties of Horizontal Axo-axonic Cells in
Stratum Oriens of the Hippocampal Ca1 Area of Rats in Vitro.
Hippocampus. 14, 232–243. • DOI: 10.1002/hipo.10170
Gentet, Luc J. – Kremer, Y. – Taniguchi,
H. – Huang, Z. J. – Staiger, J. F. – Petersen, C. C. (2012): Unique
Functional Properties of Somatostatin-Expressing Gabaergic Neurons in
Mouse Barrel Cortex. Nature Neuroscience. 15, 607–612. •
doi:10.1038/nn.3051
Jones, Edward G. (1975): Varieties and
Distribution of Non-pyramidal Cells in the Somatic Sensory Cortex of
the Squirrel Monkey. The Journal of Comparative Neurology. 160,
205–268. • DOI: 10.1002/cne.901600204
Kisvárday Zoltán F. – Adams, C. B. T. –
Smith, A. D. (1986): Synaptic Connections of Axo-axonic (Chandelier)
Cells in Human Epileptic Temporal Cortex. Neuroscience. 19, 1179–1186.
Klausberger, Thomas – Magill, P. J. –
Marton L. – Roberts, J. D. B. – Cobden, P. M. – Buzsáki G. – Somogyi
P. (2003): Brain State- and Cell Type-Specific Firing of Hippocampal
Interneurons in Vivo. Nature. 421, 844–848. • doi:10.1038/nature01374
Kittel Ágnes (2012): Örökség a jövőnek: A
Szentágothai-év nyitóeseménye Szegeden. Természet Világa. 143, 98–100.
•
WEBCÍM >
Lapray, Damien – Lasztóczi B. – Lagler M.
– Viney, T. J. – Katona L. – Valenti O. – Hartwich, K. – Borhegyi Z. –
Somogyi P. – Klausberger, T. (2012): Behavior-dependent Specialization
of Identified Hippocampal Interneurons. Nature Neuroscience. 15,
1265–1271. • doi:10.1038/nn.3176
Li, X-G. – Somogyi P. – Tepper, J. M. –
Buzsáki Gy. (1992): Axonal and Dendritic Arborization of an
Intracellularly Labeled Chandelier Cell in the Ca1 Region of Rat
Hippocampus. Experimental Brain Research. 90, 519–525. •
DOI: 10.1007/BF00230934
Lörincz Andrea – Nusser Zoltán (2008): Cell-Type-Dependent Molecular
Composition of the Axon Initial Segment. The Journal of Neuroscience.
28, 14329–14340. • doi: 10.1523/JNEUROSCI. 4833-08.2008 •
WEBCÍM >
Molnár Gábor – Oláh S. – Komlósi G. – Füle
M. – Szabadics J. – Varga C. – Barzó P. – Tamás G. (2008): Complex
Events Initiated by Individual Spikes in the Human Cerebral Cortex.
Plos Biology. 6, 1842–1849. • doi:10.1371/journal.pbio.0060222 •
WEBCÍM >
Nusser Zoltán – Sieghart, W. – Benke, D. –
Fritschy, J-M. – Somogyi P. (1996): Differential Synaptic Localization
of Two Major G-Aminobutyric Acid Type A Receptor a Subunits on
Hippocampal Pyramidal Cells. Proceedings of the National Academy of
Sciences of the USA. 3, 11939–11944. •
WEBCÍM >
Rancz Ede A. – Franks, K. M. – Schwarz, M.
K. – Pichler, B. – Schaefer, A. T. – Margrie, T. W. (2011):
Transfection via Whole-Cell Recording in Vivo: Bridging Single-Cell
Physiology, Genetics and Connectomics. Nature Neuroscience. 14,
527–532. • doi:10.1038/nn.2765
Ribak, Charles E. (1978): Aspinous and
Sparsely-Spinous Stellate Neurons in the Visual Cortex of Rats Contain
Glutamic Acid Decarboxylase. Journal of Neurocytology. 7, 461–478.
• DOI: 10.1007/BF01173991
Somogyi Péter (1977): A Specific
‚Axo-axonal’ Interneuron in the Visual Cortex of the Rat. Brain
Research. 136, 345–350. •
WEBCÍM >
Somogyi Péter (2010):
Hippocampus—Intrinsic Organisation. In: Shepherd, Gordon M. –
Grillner, Sten (eds.): Handbook of Brain Microcircuit. Oxford
University Press, Oxford, 148–164. •
WEBCÍM >
Somogyi Péter – Freund Tamás – Cowey, A.
(1982): The Axo-axonic Interneuron in the Cerebral Cortex of the Rat,
Cat and Monkey. Neuroscience. 7, 2577–2607. •
WEBCÍM >
Somogyi Péter – Nunzi, M. G. – Gorio, A. –
Smith, A. D. (1983a): A New Type of Specific Interneuron in the Monkey
Hippocampus Forming Synapses Exclusively with the Axon Initial
Segments of Pyramidal Cells. Brain Research. 259, 137–142. •
WEBCÍM >
Somogyi Péter – Smith, A. D. – Nunzi, M.
G. – Gorio, A. – Takagi, H. – Wu, J. Y. (1983b): Glutamate
Decarboxylase Immunoreactivity in the Hippocampus of the Cat.
Distribution of Immunoreactive Synaptic Terminals with Special
Reference to the Axon Initial Segment of Pyramidal Neurons. The
Journal of Neuroscience. 3, 1450–1468. •
WEBCÍM >
Somogyi Péter – Freund T. F. – Hodgson, A.
J. – Somogyi J. – Beroukas, D. – Chubb, I. W. (1985): Identified
Axo-axonic Cells Are Immunoreactive for Gaba in the Hippocampus and
Visual Cortex of the Cat. Brain Research. 332, 143–149. •
WEBCÍM >
Szabadics János – Varga C. – Molnár G. –
Oláh S. – Barzó P. – Tamás G. (2006): Excitatory Effect of Gabaergic
Axo-axonic Cells in Cortical Microcircuits. Science. 311, 233–235. •
DOI: 10.1126/science.1121325
Szentágothai János (1975): The
‚Module-Concept’ in Cerebral Cortex Architecture. Brain Research. 95,
475–496. • http://dx.doi.org/10.1016/0006-8993(75) 90122-5
Szentágothai János (1978): The Neuron
Network of the Cerebral Cortex: A Functional Interpretation. The
Ferrier Lecture, 1977. Proceedings of the Royal Society B: Biological
Sciences. 201, 219–248.
Szentágothai János – Arbib, Michael A.
(1974): Conceptual Models of Neural Organization. Neurosciences
Research Program Bulletin. 12, 305–510.
Zhu, Yinghua – Stornetta, R. l. – Zhu, J.
J. (2004): Chandelier Cells Control Excessive Cortical Excitation:
Characteristics of Whisker-Evoked Synaptic Responses of Layer 2/3
Nonpyramidal and Pyramidal Neurons. The Journal of Neuroscience. 24,
5101 – 5108. • doi: 10.1523/JNEUROSCI.0544-04.2004 •
WEBCÍM >
|
|