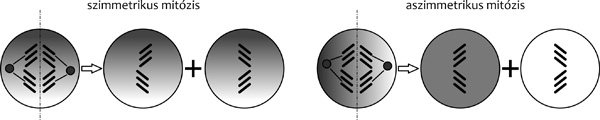
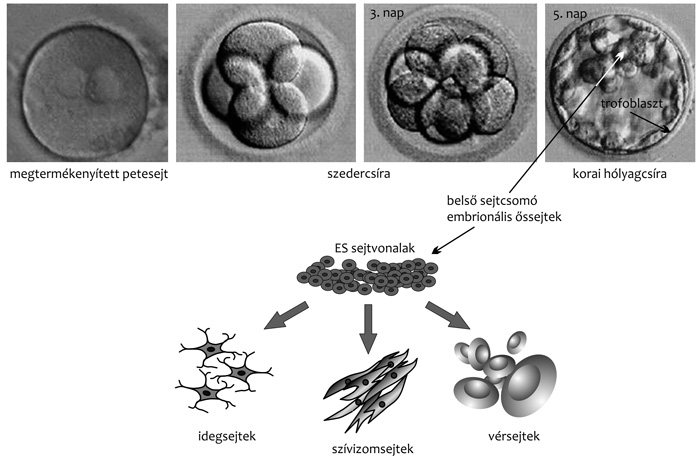
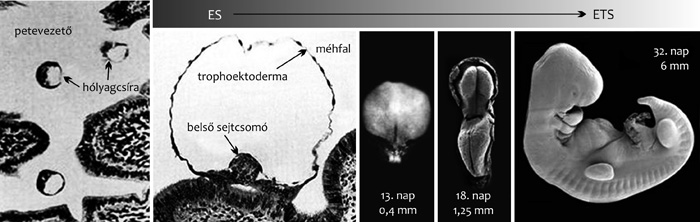
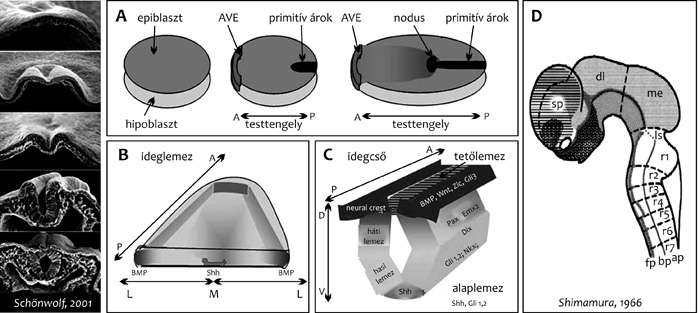
|
A fej-farok irányú pozícionálás információ mellett,
az embriólemez középvonalában (például: sonic hedgehog [Shh]; TGF-b)
és az oldalsó szegélyen (például: BMP, Wnt-faktorok) termelődő
morfogének meghatározzák a test medio-laterális tengelyét (4.
ábra B). Ez a korai pozicionális információ biztosítja a jövendő
test bilaterális szimmetriáját, és az egyes létfontosságú szervek
(például: szív, máj) egyoldali elhelyezkedését. A csővé formálódó
központi idegszövetben ez a korai medio-laterális – majd a csővé
formálódás után ventro-laterális – meghatározódás (4. ábra C)
biztosítja többek között a gerincvelői mozgató idegsejtek ventrális,
és az érző bemenetek dorzális szerveződését. A morfogén-gradiensek
és az általuk indukált génexpressziós mintázatok a fejlődő
embriótestet és test teljes anterior–poszterior tengelye mentén
végighúzódó központi idegszövet-kezdeményt (4. ábra D) egymástól
elkülönülő doménekre osztják. Az egyes domének sejtjei a normális
fejlődés során egymástól eltérő sajátságú utódokat, jellegzetes
sejtmintázatokat, azaz a test megfelelő helyén megfelelő sajátságú
differenciálódó szövetegyütteseket, szervkezdeményeket hoznak létre.
A szövet- és szervképzés időszakában az embrionális
szöveti őssejtek már jelentősen különböznek egymástól: a vér- és
érképző, vázrendszert építő vagy idegi őssejtek jellegzetes alkotó
molekuláik alapján jól elkülöníthetőek. A sejtfejlődés során az
egyes sejtféleségek azáltal különülnek el egymástól, hogy azonos
génállományukból más és más génszakaszok működnek aktívan, illetve
„hallgatnak el”. A fejlődés megfordíthatatlan folyamatait a
génállomány azon változásai okozzák, amelyek egyes génszakaszok
végleges inaktiválódásával járnak. Az ilyen változások – például a
DNS-metilációk, -deléciók – a sejtfejlődés során halmozódnak,
végdifferenciált sejtek kialakulásához vezetnek, és azt
eredményezik, hogy az ilyen sejtek környezetükből kiragadva sem
alakíthatók más sejtféleséggé.
Szöveti őssejtek a felnőtt szervezetben
A szövet- és szervépítés „látványos” folyamatai lényegében a
felnőtté válással fejeződnek be. Szöveti őssejtek azonban a test
egyes részeinek – bőr, bélcsatornahám, száj- és orrnyálkahártya,
csontvelő vérképző szigetei, előagyi neurogén zónák – germinatív
sejtcsoportjaiban fennmaradnak, és aktívan képeznek sejteket az élet
teljes időtartama alatt. Működésük nélkül a szervezet nem létezhet.
Emellett minden szervben megtalálhatók azok a nem végdifferenciált
sokszorozó sejtek, amelyek biztosítják, hogy a szervek gyorsan
átrendeződő szerkezeti elemei – kötőszövetes burkok, plasztikusan
változó kapillárisok és limfa-erek – normális működési feltételek
mellett is állandóan változzanak.
Noha minden szövetben jelen vannak őssejtek, nem
minden kifejlett szerv vagy szövet képes regenerálódni, és a
sérülések után regenerálódó szövet állaga lényegesen eltérhet az
egészségestől. A kifejlett szövetekben sem az őssejtek, sem az
azokat fogadni tudó szöveti környezet nem azonos a korai
szövetfejlődés időszakában létezővel. Ma még keveset tudunk a
felnőttkori őssejtek sajátságairól és a fejlődésükhöz, irányított
vándorlásukhoz szükséges környezeti feltételekről; jelenleg ezt a
jelentős regeneratív lehetőséget a gyógyításban alkalmazni még nem
tudjuk. Komoly kutatómunkára van szükség, hogy megértsük, hányféle
különböző típusú őssejt létezik az egyes érett szövetekben, és ezek
milyen lényeges sajátságokban térnek el egymástól. Meg kell
értenünk, hogy hogyan képesek ezek a sejtek megtartani
őssejtállapotukat, miközben a környező sejtek magasan fejlett
végdifferenciált sejtekké alakulnak. Az sem világos, vajon a
felnőttkori őssejtek az egyes szöveti-fejlődési lépéseket képviselő
őssejtek fennmaradt reprezentánsai-e, vagy a szöveti fejlődés más
folyamatában keletkeznek. Nem világos, hogy e sejtek közül melyek
képesek – ha képesek – más szövetek sejttípusait is létrehozni.
Ez utóbbi kérdéseket behatóan vizsgálják a
felnőttkori testi sejtek „visszaprogramozásával” és/vagy
„transzdifferenciálódásával” foglalkozó kutatások.
Szöveti őssejtek „visszaprogramozása”
és „transzdifferenciálódása”
A felnőtt emberi szöveti őssejtek felbecsülhetetlen forrást ígérnek
a saját sejttel való jövőbeli klinikai sejtpótláshoz, az egyedi
genetikai/sejtbiológiai sajátosságok diagnosztikus elemzéséhez és az
egyedre szabott gyógyszerhatások vizsgálataihoz. A testből nyert, in
vitro fenntartható sejtek csakis az osztódóképességüket megőrző, nem
végdifferenciált szöveti sejtek lehetnek. Ezek között a szöveti
ős-/sokszorozó sejtek között kereshetők azok, amelyekben előidézhető
egy kevéssé differenciált, többféle sejtfejlődésre képes,
pluripotens sejtállapot.
Embrionális őssejtekre jellemző gének aktiválásával
indukálni lehet embrionális őssejt- (ES) sajátságokat a felnőtt – de
nem végdifferenciált – testi sejtek egész során.
Az ún. őssejtgének aktiválását el lehet érni e gének sejtekbe való
közvetlen bevitelével (Takahashi – Yamanaka, 2006), vagy a sejtek
génszabályozó fehérjehálózatának kis molekulákkal vagy növekedési
faktorokkal (Hussein – Nagy 2012; Liang – Zhang, 2013) való
szabályozásával. Ez a „visszaprogramozás” azonban csak osztódóképes,
azaz nem végdifferenciált sejtekben (1. táblázat) működik.
A visszaprogramozás eredményeként ES-sejtekhez
hasonló, elkötelezetlen sejteket állítanak elő; ma az indukált
pluripotens sejtek minőségét is az ES-kritériumok szerint ítélik
meg. Ez azonban azt jelenti, hogy szöveti fejlődésüket, és főként
egy lehetséges jövőbeni klinikai alkalmazást tekintve hasonló
problémákkal szembesülnek a kutatók, mint az ES-sejtek esetén:
teljesen elkötelezetlen őssejtek közvetlenül nem használhatók
szövetképző sejtek modellezésére vagy pótlására. A gyakorlatban
szélesen alkalmazható visszaprogramozás távlati – ma még
futurisztikus – célja az lenne, hogy olyan szöveti őssejteket hozzon
létre, amelyek az adott szövet alakításához az éppen szükséges
sejtfejlődési állapotot képviselik. Ehhez azonban az egyes szöveti
őssejtek szövetképző potenciálját, finom sajátosságait és környezeti
igényeit kellene ismerni. A rohamos tudományos fejlődés ellenére
ezekre az adatokra ma még várni kell. A megoldandó kérdésekről és a
már elért eredményekről részletesen szólnak e kötet tanulmányai.
Kulcsszavak: szöveti fejlődés, regeneráció,
sejttípus-elköteleződés, végdifferenciált sejtek, testtengelyek
IRODALOM
Hussein, S. M. - Nagy A. (2012): Progress
Made in the Reprogramming Field: New Factors, New Strategies and a
New Outlook. Current Opinion in Genetics & Development. 22, 435-43.
Liang, G. – Zhang, Y. (2013): Embryonic
Stem Cell and Induced Pluripotent Stem Cell: An Epigenetic
Perspective. Cell Research. 23, 49–69.
Schoenwolf, G. C. (2001): Cutting, Pasting
and Painting: Experimental Embryology and Neural Development. Nature
Reviews Neuroscience. 2, 11, 763–771.
Shimamura, K. – Hartigan, D. J. –
Martinez, S. – Puelles, L. – Rubenstein, J. L. R. (1995):
Longitudinal Organization of the Anterior Neural Plate and Neural
Tube. Development. 121, 3923–3933.
Stadtfeld, M. – Hochedlinger, K. (2010):
Induced Pluripotency: History, Mechanisms, and Applications. Genes &
Development. 24, 2239–2263.
Takahashi, K. – Yamanaka, S. (2006):
Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult
Fibroblast Cultures by Defined Factors. Cell. 126, 663–676.
Yamanaka, S. (2012): Induced Pluripotent
Stem Cells: Past, Present, and Future. Cell Stem Cell. 10, 678–684.
|