|
genetikai szempontból is sokkal „mozaikosabbak”
lehetnek, mint azt eddig gondoltuk. Régóta ismert volt például, hogy
az emberi genom rengeteg mobilis genetikai elemet (transzpozont)
tartalmaz, ám ezekre sokáig mint többségükben nem aktív genetikai
parazitákra gondoltunk. Kiderült azonban, hogy bizonyos
retrotranszpozonok az ősivarsejteken (a „csíravonal” sejtjein) kívül
adott szöveti sejtek korai előalakjaiban, így például idegi
előalakokban (neuronális progenitor sejtekben) is aktívak lehetnek.
Márpedig így a fejlődés során heterogén sejtpopulációk jöhetnek
létre, ami végső soron szomatikus mozaicizmushoz, vagyis a hasonló
típusú testi sejtek genetikai állományának apró különbségeihez vezet
(Singer et al., 2010). Nem tudjuk, hogy ez a folyamat mennyire
érinti például az IPS-sejtek előállításához gyakran használt
kötőszöveti sejteket, de az biztos, hogy az IPS-sejtekből
differenciációval létrehozott testi sejtek akár emiatt is
különbözhetnek a kiindulási „gazdaszervezet” hasonló testi
sejtjeitől (Abyzov et al., 2012). Hasonló problémát jelentenek a
testi sejtjeinkben az évek során spontán felhalmozódó mutációk is,
hiszen egy idősebb ember kötőszöveti sejtjei statisztikailag is
jóval több genetikai eltérést tartalmazhatnak, mint egy fiatalé, így
egy esetleges szervpótlás céljából az IPS-sejtek előállításához
kinyert sejtek genetikai szempontból nem feltétlenül jelentik az
„ideális” forrást. Ezek a megfigyelések természetesen nem
kérdőjelezik meg az IPS-technológia létjogosultságát, de felhívják a
figyelmet olyan új vizsgálati szempontokra, amelyekkel akár az így
létrehozott betegségmodellek reprezentatív voltát, akár a
szövetpótlás során visszaültetendő sejtek „minőségét” is fontos lesz
majd a jövőben ellenőrizni.
Szöveti sejtek egymást közt –
a transzdifferenciáció folyamata
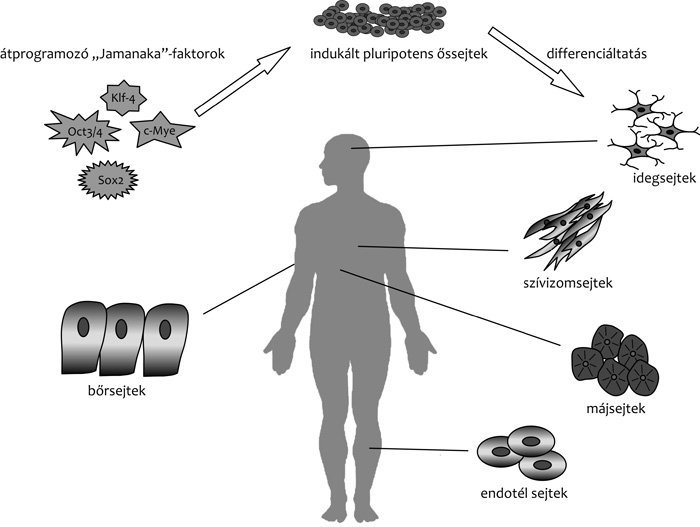
A Jamanaka Sinja munkacsoportja által először elírt IPS-technológia
kétségkívül forradalmasította a sejtek differenciációjáról alkotott
képünket, és a korábban irreverzibilisnek gondolt folyamatokról
kiderült, hogy bizonyos körülmények között igenis van lehetőség a
testi sejtek egy korábbi állapotba történő visszaállítására.
Mindezek kapcsán azonban felmerült az a gondolat is, hogy genetikai
szempontból hátha van közvetlenebb „átjárás” a testi sejtek között,
vagyis lehet-e horizontális irányban is átszelni a magaslatokat a
korábban már említett Waddington-tájképen. A modern
sejtdifferenciációs kísérletek pedig igazolni látszanak ezeket a
feltételezéseket, ugyanis egyre több esetben képesek a kutatók a
transzdifferenciáció segítségével adott testi sejtekből más
típusúakat előállítani. Sikerült például kötőszövetek közvetlen
átprogramozásával szívizomsejteket előállítani (Qian et al., 2012;
Song et al., 2012), vagy akár emberi vizeletből származó sejtekből
idegsejt előalakokat létrehozni (Wang et al., 2012). A módszer
minden esetben azon alapul, hogy a legtöbb testi sejt egyedi
génexpressziós mintázattal rendelkezik, de sok esetben egy jól
definiálható kisebb számú géncsoport (jellemzően transzkripciós
faktorok) erőteljes kifejeződése alakítja ki az adott sejtre
jellemző fenotípust, vagyis határozza meg a sejtek identitását.
Innen származik az ötlet, hogy akkor ezen meghatározott faktorok
kívülről történő „erőltetett” expressziójával a sejtek identitását
esetleg meg lehet változtatni. Természetesen vannak olyan speciális
alakú és működésű végdifferenciált sejtek (akár bizonyos csontsejtek
vagy idegsejtek), amelyek esetében kevésbé várható egy ilyen gyors
átalakulás, a sokkal plasztikusabb kötőszöveti sejtek azonban akár
erre is alkalmasak lehetnek (Sancho-Martinez et al., 2012).
De miért merül fel egyáltalán az igény arra, hogy
átprogramozzunk adott sejteket más típusúakká, amikor embrionális
őssejtekből vagy IPS-sejtekből elvileg akármilyen sejttípus
előállítható? A válasz a gyorsaságban, hatékonyságban és a
potenciális veszélyek kiiktatásában rejlik. Ha ugyanis rendelkezésre
áll a megfelelő forrássejt, akkor a transzdifferenciáció
hatékonysága összemérhető az embrionális őssejtekből történő
differenciációval, főleg ha figyelembe vesszük, hogy az utóbbi
esetben bizonyos típusú sejtek vagy szövetek létrehozására és
szelektív kiválogatására sok esetben még nincs kidolgozott és
megbízható protokoll. Ha ehhez még hozzávesszük az IPS-sejtek
előállítását, ellenőrzését, és az ezen sejtekből kiinduló
differenciációt, akkor az átprogramozás lehetősége – amennyiben az
adott helyzetben tényleg alkalmazható – mindenképpen vonzó
alternatívának kínálkozik. Egy eklatáns példa erre a szívizomsejtek
létrehozása a környező szövetekben található egyéb típusú sejtekből,
amelyre jellemzően a szívizomsejteket ért károsodás (például
infarktus) utáni helyzetben van szükség, ahol a hatékonyság és az
időfaktor döntő jelentőségű lehet. Szívizomsejtekre jellemző
transzkripciós faktorok (ilyenek a GATA4, a MEF2C, vagy a TBX5)
túlzott expressziójával több munkacsoportnak sikerült az előbbiekben
leírt módon szívizomsejteket előállítani, és több esetben
bizonyították, hogy az így létrehozott sejteket állati szívbe
visszaültetve azok képesek voltak a megfelelő funkciót ellátni (Qian
et al., 2012).
Az átprogramozás sikeressége nagyban függ attól, hogy találunk-e
olyan „mester regulátor” géneket, amelyek a kívánt szövet
expressziós profiljának meghatározott domináns elemeit jelentik, és
amelyek fokozott expressziójától várható, hogy elősegítik a sejt
identitásának konverzióját. A szívizomsejteken kívül eddig
bizonyítottan májsejteket, illetve különböző típusú idegsejteket
sikerült már ilyen módszerrel előállítani, mert ezen sejttípusokban
már korábban azonosítottak a sejtek fenotípusának kialakításában
szerepet játszó domináns géneket (Sancho-Martinez et al., 2012).
Természetesen akárcsak az IPS-technológia esetében, ehhez a
módszerhez is jól jöhetnek olyan kis molekulák (vagy akár adott
fehérjéket kódoló gének), amelyek a sejtek epigenetikai memóriájának
átalakításában vagy letörlésében szerepet játszanak, befolyásolva
például a DNS metilációs mintázatát, vagy a DNS köré csavarodó
hisztonfehérjék kovalens módosításait. Ezek alkalmazása a
hatékonyság növelése mellett azonban az őssejti állapothoz közelítve
növelheti a rákos fenotípus kialakulásának veszélyét.
Kétségtelen azonban, hogy a transzdifferenciáció
nagyon ígéretes technológiának tűnik nemcsak a regeneratív orvoslás
területén, hanem adott modellsejtekre épülő gyógyszerhatástani és
toxikológiai szűrővizsgálatok során is. Nem kérdés, hogy a májsejtek
például nagyon kedvelt farmakológiai célpontok, hiszen
használatukkal jól modellezhető például új készítmények metabolikus
lebontásának a szervezeten belüli útvonala. Emiatt komoly piaci
igény van arra, hogy ilyen típusú sejteket nagy mennyiségben,
hatékony módon és viszonylag rövid idő alatt elő lehessen állítani,
és a szöveti átprogramozás folyamata kétségkívül megoldást jelenthet
erre a problémára. Az ígéretes kezdeti eredmények dacára azonban
ezen technológia esetében sem szabad elfeledkezni a kötelező
minőségellenőrzési pontokról és a potenciális veszélyforrások
lehetőség szerinti elkerüléséről. Fontos szempont például, hogy sok
transzkripciós faktor bizonyos daganatos sejtekben is kifejeződik,
így az IPS-technológiához hasonlóan a transzdifferenciáció esetében
is szem előtt kell tartani a rákkeltő hatást mint potenciális
veszélyforrást. Nem elhanyagolható szempont ebben az esetben sem a
szomatikus mozaikosság, beleértve az idők során a sejtekben
felhalmozódott mutációkat, hiszen ezek is komolyan befolyásolhatják
az átprogramozás során létrehozott sejtek funkcióját.
A sejtprogramozás távlatai
Köszönhetően a molekuláris biológiai vizsgálómódszerek robbanásszerű
fejlődésének, az elmúlt két évtizedben a biológiai folyamatokról
alkotott tudásunk eddig nem látott óriási mértékű gyarapodáson ment
keresztül, amely talán a XX. század elején a fizikában bekövetkezett
forradalmi változásaihoz hasonlítható. A sejtek differenciációs
folyamatainak alaposabb, molekuláris genetikai szintű szabályozási
folyamatainak megismerése orvosbiológiai szempontból talán az egyik
legígéretesebb eredménynek számít, amit leginkább a 2012. évi orvosi
és élettani Nobel-díj fémjelez, amelyet az „érett sejtek pluripotens
állapotú sejtekké történő újraprogramozásának felfedezéséért”
ítéltek oda. Az IPS-technológiával, illetve a transzdifferenciáció
segítségével létrehozott sejtek felhasználása mind a regeneratív
orvoslás, mind pedig a gyógyszerkutatásban használatos sejtes
modellrendszerek területén valóban forradalmi újdonságnak számít.
Mindezen technológiák alaposabb feltérképezése kapcsán felmerülnek
olyan biológiai és orvosi problémák, amelyek az így létrehozott
sejtek biztonságos felhasználhatóságát jelenleg még korlátozzák,
ugyanakkor a sejtbiológiai és a molekuláris biológiai módszerek
fejlődésével ezek az akadályok a jövőben várhatóan elháríthatóak
lesznek. Fel kell azonban készülni arra, hogy ezen új biológiai
megközelítések kapcsán komoly etikai problémákkal is szembe kell
majd nézni, melyek a modern ember számára talán még nagyobb kihívást
jelentenek, mint a hatalmas információáradatból származó ismeretek
tudományos szintű feldolgozása. Emiatt nagyon fontos a tudományos
eredmények folyamatos és közérthető kommunikációja a társadalom
felé, és remélhetőleg a jövőben sikerül majd olyan megbízható és
átlátható ellenőrzési rendszereket kidolgozni, amelyek segítségével
a közvélemény számára is megnyugtató módon lehet ezeket a valóban
ígéretes új technológiákat a gyógyítás szolgálatába állítani.
Kulcsszavak: őssejtek, indukált pluripotens sejtek, genetikai
visszaprogramozás, átprogramozás, transzkripciós faktorok
IRODALOM
Abyzov, A. – Mariani, J. – Palejev, D. –
Zhang, Y. – Haney, M. S. – Tomasini, L. – Ferrandino, A. F. –
Rosenberg Belmaker, L. A. – Szekely, A. – Wilson, M. – Kocabas, A. –
Calixto, N. E. – Grigorenko, E. I. – Huttner, A. – Chawarska, K. –
Weissman, S. – Urban, A. E. – Gerstein, M. – Vaccarino, F. M.
(2012): Somatic Copy Number Mosaicism in Human Skin Revealed by
Induced Pluripotent Stem Cells. Nature. 492, 7429, 438–442.
Ivics Z. – Hackett, P. B. – Plasterk, R.
H. – Izsvák Zs. (1997): Molecular Reconstruction of Sleeping Beauty,
A Tc1-Like Transposon from Fish, and Its Transposition in Human
Cells. Cell. 91, 4, 501–510.
Maury, Y. – Gauthier, M. – Peschanski, M.
– Martinat, C. (2012): Human Pluripotent Stem Cells for Disease
Modelling and Drug Screening. Bioessays: News and Reviews in
Molecular, Cellular and Developmental Biology. 34, 1, 61–71.
Qian, L. – Huang, Y. – Spencer, C. I. –
Foley, A. – Vedantham, V. – Liu, L. – Conway, S. J. – Fu, J. D. –
Srivastava, D. (2012): In Vivo Reprogramming of Murine Cardiac
Fibroblasts into Induced Cardiomyocytes. Nature. 485, 7400, 593–598.
Robinton, D. A. – Daley, G. Q. (2012): The
Promise of Induced Pluripotent Stem Cells in Research and Therapy.
Nature. 481, 7381, 295–305.
Sancho-Martinez, I. – Baek, S. H. –
Izpisua Belmonte, J. C. (2012): Lineage Conversion Methodologies
Meet the Reprogramming Toolbox. Nature Cell Biology. 14, 9, 892–899.
Silva, J. – Barrandon, O. – Nichols, J. –
Kawaguchi, J. – Theunissen, T. W. – Smith, A. (2008): Promotion of
Reprogramming to Ground State Pluripotency by Signal Inhibition.
Plos Biology. 6, 10, E253.
Singer, T. – McConnell, M. J. – Marchetto,
M. C. – Coufal, N. G. – Gage, F. H. (2010): Line-1 Retrotransposons:
Mediators of Somatic Variation in Neuronal Genomes? Trends in
Neurosciences. 33, 8, 345–354.
Song, K. – Nam, Y. J. – Luo, X. – Qi, X. –
Tan, W. – Huang, G. N. – Acharya, A. – Smith, C. L. – Tallquist, M.
D. – Neilson, E. G. – Hill, J. A. – Bassel-Duby, R. – Olson, E. N.
(2012): Heart Repair by Reprogramming Non-myocytes with Cardiac
Transcription Factors. Nature. 485, 7400, 599–604.
Takahashi, K. – Tanabe, K. – Ohnuki, M. –
Narita, M. – Ichisaka, T. – Tomoda, K . – Yamanaka, S. (2007):
Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by
Defined Factors. Cell. 131, 5, 861–872.
Takahashi, K. – Yamanaka, S. (2006):
Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult
Fibroblast Cultures by Defined Factors. Cell. 126, 4, 663–676.
Wang, L. – Huang, W. – Su, H. – Xue, Y. –
Su, Z. – Liao, B. – Wang, H. – Bao, X. – Qin, D. – He, J. – Wu, W. –
So, K. F. – Pan, G. – Pei, D. (2012): Generation of Integration-free
Neural Progenitor Cells from Cells in Human Urine. Nature Methods.
10, 1, 84–89.
Woltjen, K. – Michael, I. P. – Mohseni, P.
– Desai, R. – Mileikovsky, M. – Hamalainen, R. – Cowling, R. – Wang,
W. – Liu, P. – Gertsenstein, M. – Kaji, K. – Sung, H. K. – Nagy A.
(2009): PiggyBac Transposition Reprograms Fibroblasts to Induced
Pluripotent Stem Cells. Nature. 458, 7239, 766–770.
|