|
|
Különleges adenovírusok felfedezése
hazai szarvasmarhákban
A későbbi akadémikus, Bartha Adorján (1923-1996) állatorvos
virológus az 1960-as évek elején a hazánkban akkor még újdonságnak
számító, egyrétegű, primer sejttenyészetek használatával kezdte
vizsgálni a háziállatok vírusait, amelyeket korábban csak állatról
állatra történő oltással lehetett tanulmányozni. Az újonnan
bevezetett módszerrel számos, korábban ismeretlen vírust, közöttük
adeno- és herpeszvírusokat sikerült kimutatnia szarvasmarhákból, és
elkülöníteni (izolálni) további vizsgálatokhoz (Bartha, 1969). Az
adenovírusok közepes méretű, duplaszálú DNS-t tartalmazó vírusok,
amelyek emberben és gerinces állatokban fordulnak elő. Rendszerint
ártalmatlanok, vagy csak könnyű lefolyású, spontán gyógyuló
betegséget okoznak, de immunhiányos állapotú gazdában súlyos
megbetegedések, esetenként halálos kimenetelű fertőzések is
kialakulhatnak (Benkő, 2008). Egyes szerzők a fejlett világ
országaiban manapság szinte járványszerűen terjedő, kóros elhízás
hátterében bizonyos adenovírusok oktani szerepét feltételezik, de
ezt igazolni egyelőre még nem sikerült.
Barthának szintén volt egy, a maga idejében
hihetetlennek tűnő felismerése, amelynek helyességét a későbbi
vizsgálatok világosan bizonyították. Megfigyelte, hogy az ismert
szarvasmarha-adenovírus törzsek nagyobb része, köztük három magyar
izolátum, szerológiai tesztek alapján jól elkülönül az emberből,
illetve más, emlősállatokból (például: kutyából, juhból, lóból)
kimutatott adenovírusoktól, és több biológiai tulajdonságban is
markánsan eltér azoktól. A szarvasmarha-adenovírusok e csoportjára
jellemző volt az általuk okozott magzárványok jellegzetes
morfológiája, fokozott hőtűrő képességük, és az, hogy
replikációjukhoz primer, fibroblaszt típusú sejttenyészetet
igényeltek. Noha a vírusok rendszertana ekkor még gyermekcipőben
járt, az emberben (emlősökben), illetve a madarakban talált
adenovírusokat már két külön csoportba, későbbi nemzetségbe
(Mastadenovirus vagy Aviadenovirus genusba) sorolták. Bartha az
általa talált, különleges tulajdonságokkal jellemezhető típusokat
annyira eltérőnek vélte, hogy javaslatot tett külön alcsoportba
sorolásukra, sőt az összes többi emlős adenovírusától való
elkülönítésükre (Bartha, 1969). Ezt a javaslatot azonban nem túl
meglepő módon, a szinte kizárólag nyugati kutatókból álló
rendszertani bizottság csak feltételesen, „további bizonyítékok”
függvényében fogadta el. Fenntartásaikban nyilván szerepet játszott
a vasfüggönyön túlról származó minden tudományos eredménnyel
szembeni természetes gyanakvás, de a kételyeket fokozhatta az is,
hogy a legalaposabban tanulmányozott gazdában, az emberben ilyen
különleges adenovírusokat sosem találtak. Az adott kor technikájával
valóban nem lehetett választ adni arra, hogy a megfigyelt eltérések
mértéke csakugyan elegendő-e egy új genus létrehozásához. A közeli
vírusok közötti különbségek szabatosan mérhetőek voltak a
vírus-neutralizációs próbában adott keresztreakciók számszerűsített
mértékével. A keresztreakció hiánya azonban értelemszerűen nem
szolgáltathatott kvantitatív adatokat. Arra viszont fény derült,
hogy miért éppen egy szegény ország kutatója találja egymás után
ezeket az új vírusokat. Eleinte ugyanis Barthának és a többi magyar
virológusnak nem volt lehetősége a nyugati országokban már
fokozatosan terjedő, „halhatatlanná tett” (elméletileg a
végtelenségig szaporítható, azaz passzálható) sejtvonalakhoz
hozzájutni. A hazai kutatók a hizlalásra szánt hím állatok
ivartalanítása során eltávolított heréből vagy a vágóhidakról
beszerzett, más belső szervekből készítettek mindig friss, primer
sejttenyészeteket, és a különleges (2. alcsoportba sorolt)
szarvasmarha-adenovírusokat csak ezeken lehetett izolálni, de
gyakran spontán sejtkárosodás is kialakult, ha a donor állat eleve
fertőzött volt adenovírussal.
A vírusok összehasonlító
genetikai vizsgálatának kezdetei
A vírusok közötti különbségek mérhetőségével, illetve
számszerűsítésével a molekuláris módszerek hazánkba érkezéséig
kellett várni. Bartha (akkor még) fiatal munkatársa, Benkő Mária
először a laboratórium törzsgyűjteményének vírusaiból kivont DNS
restrikciós enzimekkel való kezelése után kapott genomtöredékek
méreteinek összehasonlítása alapján (Benkő et al., 1988), majd
később DNS-hibridizációs technikával már ki tudta mutatni, hogy az
összes emlős adenovírusa jobban hasonlít egymáshoz, mint a
szarvasmarha-adenovírusok két alcsoportjának tagjai, noha ezek
azonos gazdából származnak (Benkő et al., 1990). Végül a
DNS-szekvenálás és az ennek nyomán terjedő
filogenetikai számítások (amit állati adenovírusok
összehasonlítására elsőként magyar kutatók alkalmaztak) meggyőző és
kvantitatív módon bizonyították, hogy az evolúciós távolság kellően
nagy ahhoz, hogy ezek a különleges szarvasmarha-adenovírusok egy új
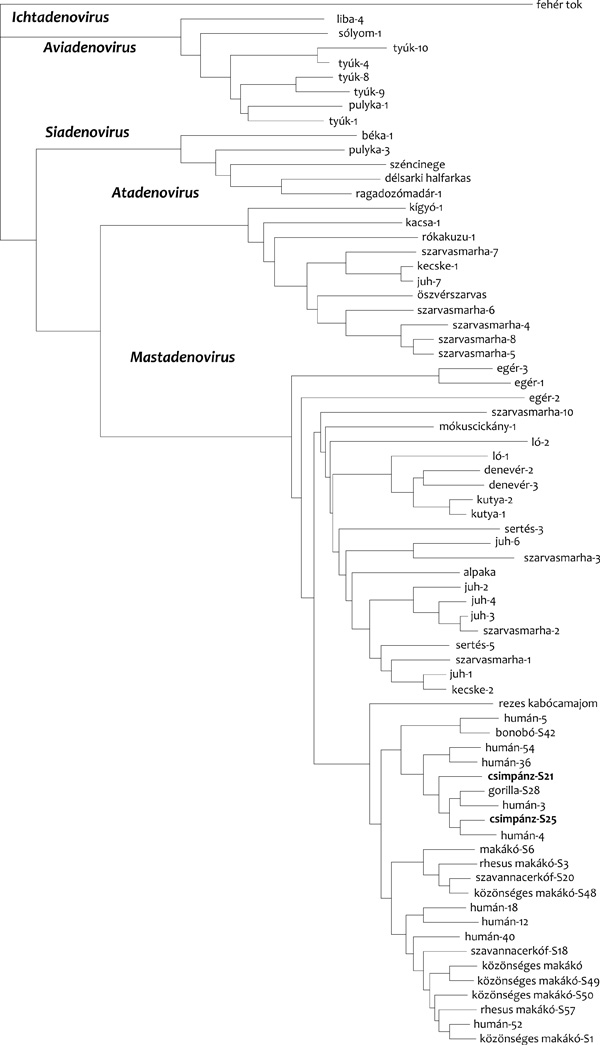
nemzetségbe kerüljenek (1. ábra)
(Harrach et al., 1997). Ráadásul egy roppant meglepő felfedezés is
történt, amennyiben a tyúkokban és libákban jellegzetes betegséget
okozó adenovírus (Ivanics et al., 2001), hivatalos nevén
kacsa-adenovírus 1, állatorvosi nevén a tojáshozam-csökkenés
szindróma (EDS) vírusa, távoli gazda-eredete ellenére is rokonnak
bizonyult e különleges szarvasmarha-adenovírusokkal (Harrach et al.,
1997). Kiderült továbbá, hogy egy időközben Ausztráliában
felfedezett és analizált, hasonlóan különleges juh-adenovírus is
ebbe az új genusba sorolható (Harrach et al., 1997; Benkő – Harrach,
1998).
De mi lehet a magyarázata annak, hogy a
meglehetősen távoli evolúciós ágon fejlődő madaraknak és kérődzőknek
ennyire hasonló adenovírusaik is vannak, amelyek ugyanakkor
jelentősen különböznek a madarakban (főként baromfiban), illetve az
emlősökben általában talált, az egymástól is élesen elkülönülő
Aviadenovirus vagy Mastadenovirus genusba sorolt vírusoktól? Ekkor
született megint egy merész (és sokáig bizarrnak tekintett) magyar
gondolat. Tételezzük fel, hogy ezek a különleges adenovírusok egy
harmadik gerinces osztály tagjaival végig együtt fejlődött
leszármazási vonalat képviselnek, és azokba az állatfajokba, amelyek
képviselőiben eddig felismertük e vírusokat, gazdaváltás útján
kerültek!
Javaslat új vírusnemzetségek alapítására
a rendhagyó adenovírusok besorolásához
A különleges adenovírusok lehetséges eredete után kutatva
meghatároztuk és elemeztük az egyetlen ismert béka-adenovírus
genomjának teljes szekvenciáját. A munka meglepő eredményt hozott,
ugyanis kiderült, hogy a békából származó adenovírus egy másik,
„rendhagyó” madár-adenovírussal, a 3-as típusú pulyka-adenovírussal,
állatorvosi nevén a pulykák vérzéses bélgyulladásának (THE)
vírusával áll nagyon közeli rokonságban (Davison et al., 2000). Az
újonnan felismert vírusnemzetségnek, amelyet akkori hipotézisünk
szerint a kétéltűekkel együtt fejlődött vonalnak gondoltuk, a
Siadenovirus nevet adtuk, utalva a csakis ezeknek az adenovírusoknak
a genomjában kimutatott, szialidázszerű gén jelenlétére. A
filogenetikai számítások során határozottan megkülönböztethető
csoportokon belül az egyes vírusok génállománya is az adott
csoportra jellemző volt (Davison et al., 2003).
Így tehát már négy jól elkülönülő vírusvonal
létezését tártuk fel, de még mindig nem tudtuk megmagyarázni a
különleges szarvasmarha/juh/tyúk/kacsa-adenovírus-vonal eredetét.
Ezért feltételeztük, hogy a kérdéses kérődző/madár-vírusok,
amelyeket genomjuk alacsony G+C-tartalma miatt AT-gazdag, azaz
atadenovírusoknak neveztünk el (Benkő – Harrach, 1998), esetleg a
hüllők adenovírusai lehetnek (Harrach, 2000). Az akkoriban a világon
egyetlen kígyó-adenovírus izolátum megszerzése Németországból
hosszas levelezést és éveket vett igénybe. Közben tudomást
szereztünk egy feltételezett hal-adenovírus izolátum létezéséről is
az Egyesült Államokban. Végül mindkét vírust elszaporítva a
rendelkezésünkre bocsátották, de azok koncentrációja (titere) nagyon
alacsony lehetett, mert a szokásos módszerekkel nehezen voltak
vizsgálhatók. Az első szekvenciaadatokat PCR (polimeráz-láncreakció)
segítségével sikerült kinyerünk (Benkő et al., 2002). A
hal-adenovírus egy teljesen új, immár ötödik adenovírus-vonalnak
bizonyult (Kovács et al., 2003). Ugyanakkor óriási megelégedéssel
nyugtáztuk, hogy a kígyó-adenovírus a törzsfákon valóban az
atadenovírusok csoportjába került (1. ábra). A teljes
szekvencia meghatározása és elemzése bizonyította, hogy a
kígyó-adenovírus genomszerveződése alapján is az Atadenovirus
nemzetségbe sorolható (Farkas et al., 2008). Amerikai kollégákkal
további hüllő- (kígyó- és gyík-) adenovírusokról bizonyítottuk be,
hogy azok szintén az atadenovírusokkal közös, közeli őstől
származnak (Wellehan et al., 2004). Német laboratóriumban
vendégkutatóként dolgozó magyar diákjaink németországi gyík- és
kígyótenyészetekben mutattak ki atadenovírusos fertőzöttséget (Papp
et al., 2009). Később az Egyesült Államokban, Új-Zélandon és
hazánkban is újabb atadenovírusok jelenlétét tárták fel különféle
gyíkokban és kígyókban, és azóta is folyamatosan nő azoknak az
eredményeknek a száma, amelyek szerint ez a leszármazási vonal a
pikkelyes hüllők saját, velük végig együtt fejlődő adenovírusainak
felel meg.
Érdekes módon azonban a pikkelyes hüllőkben
kimutatott atadenovírusok DNS-ének A+T-tartalmát nem magasnak, hanem
általában kiegyensúlyozottnak találtuk. Úgy tűnik, hogy a magas
A+T-arány a gazdaváltással lehet összefüggésben. Erre vonatkozóan
egyelőre semmi bizonyítható feltételezésünk nincs, de tény, hogy
például a macska AIDS kórokozója (feline immundeficiency virus)
esetében megfigyelték, hogy idegen gazdában (nevezetesen pumában) a
vírus genomjában gyakori A→G mutáció következik be. Természetesen
ezt a rendkívül variábilis RNS-vírusoknál megfigyelt jelenséget nem
lehet a komplex genomszerveződésű, duplaszálú DNS-vírusokra
közvetlenül vonatkoztatni, de a párhuzam elgondolkodtató, és a
jelenség feltétlenül további vizsgálatra érdemes. Mindenesetre a
nemzetség Atadenovirus neve így ma már kevésbé tetszik találónak,
mert valójában csak a vírusvonal elsőként felismert és jellemzett
tagjaira érvényes. A vírusrendszertanban helyesen alkalmazott
szabály szerint azonban ilyen esetekben is célszerűbb a már
elfogadott név megtartása. Hasonló helyzetet találunk más
víruscsaládok esetében is, így például a parvovírusok Dependovirus
nemzetségének ma már több olyan tagját is ismerjük, amely a genus
nevével ellentétben önálló, azaz képes a helpervírus nélküli
replikációra.
Annak ellenére, hogy adenovírusok jelenléte a
pikkelyes hüllőkben általában gyakori, a többi hüllőrend tagjaiban
egyelőre nem sikerült atadenovírust kimutatni. Ezért jelenlegi
álláspontunk szerint az Atadenovirus nemzetség a pikkelyes hüllőkkel
együtt fejlődött adenovírus leszármazási vonal lehet, és
elképzelhető, hogy a hüllők többi rendjébe tartozó állatokban
(krokodilok, teknősök) esetleg más, eddig ismeretlen leszármazási
vonalhoz tartozó adenovírusok fordulnak elő.
Régebbi és újabb gazdaváltások feltételezése
Az Egyesült Államokban nemrégiben érdekes felfedezés született az
országba illegálisan becsempészett és elkobzott teknősök tömeges
megbetegedését követően. A beteg, illetve elhullott egyedekben PCR
segítségével egy új siadenovírus DNS-ét tudták kimutatni, amelynek a
békából és pulykából származó siadenovírusokhoz hasonlóan szintén
magas A+T-tartalma volt. Az elkobzott teknősöket megfelelő
karanténozás nélkül, több állatkertben, illetve rezervátumban
helyezték el, és ezek némelyikében a későbbiekben egyéb teknősfajok
képviselői is megbetegedtek. Ezekből a tapasztalatokból arra
következtethetünk, hogy az újonnan felismert siadenovírus eredeti
gazdája nem a teknősök között keresendő, de a béka-siadenovírus
magas A+T-tartalma a kétéltű-eredetet is cáfolja. Nem találtunk
további siadenovírusokat kétéltűekben, noha célzottan vizsgáltunk
számos békából és farkos kétéltűből származó mintát. Ugyanakkor,
egyre gyarapszik a vad- és egzotikus (gyakran kedvencként tartott)
madarakban felismert új siadenovírusok száma. A korszerű rendszertan
szerint a madarak is hüllőknek tekintendők, tehát valószínűbb, hogy
a siadenovírusok is eredetileg egy ősibb hüllővonalban alakultak ki,
és ezért viszonylag könnyebben történhet gazdaváltásuk teknősökre és
madarakra.
Továbbra is úgy véljük, hogy az aviadenovírusok a
madarakkal folyamatosan együtt fejlődő adenovírusok, és ennek
bizonyítékául gondoljuk, hogy szinte minden alaposan vizsgált
madárfaj képviselőiben ki tudjuk mutatni a fajjal feltehetően együtt
kialakult, specifikus aviadenovírust. Jóval ritkábban és kevesebb
madárfajban fordulnak elő siadenovírusok, és ezek általában erősebb
kórokozó hatást mutatnak, mint az aviadenovírusok. Feltételezésünk
szerint a fokozott megbetegítő képesség aránylag közeli
gazdaváltásra utal. Úgy tűnik, hogy a madarakban előforduló harmadik
adenovírus nemzetség tagjai, az atadenovírusok természetes
körülmények között szinte csak az énekesmadarakban, kivételes
esetekben a zsúfoltan tartott víziszárnyasokban, baromfiban és
tenyésztett papagájokban bukkannak fel. Mint korábban kifejtettem,
az atadenovírusok pikkelyes hüllő eredetét ma már bizonyítottnak
tekintjük, ezért választottam a jelen összefoglaló ismertetésnek a
fenti, kissé bizarr címet. A házityúkban valóban előfordulhatnak
kígyóktól (vagy más pikkelyes hüllőktől) eredő adenovírusok. E
vírusok madarakra
|
|
|
történő gazdaváltásának időpontját azonban nehéz
megbecsülni. Elképzelhető, hogy nagyjából az után történhetett, hogy
az énekesmadarak (Passeriformes) ága elkülönült a többi madárétól,
hiszen az előbbiekben gyakran megtalálhatók és változatosak, míg az
utóbbiakban (baromfi, bizonyos vízimadarak) csak ritkán, és
rendszerint ugyanaz az atadenovírus fordul elő. Ez az előbbitől
teljesen független, a legújabb időkben való gazdaváltásra utal. A
házityúkban általában a kacsa-adenovírus 1-es izolátumra nagyon
hasonlító változatokat lehet csak kimutatni. A vírus által okozott
tojáshozam-csökkenéssel járó kórkép szinte azonos időben (1976-tól
kezdve) jelentkezett világszerte, és nem zárható ki, hogy „oltási
baleset” eredményeként, valamilyen általánosan használt vakcina
közvetítésével terjedt el mindenütt. A pikkelyes hüllőkről az
emlősökre (kérődzőkre és a közönséges rókakuzura) való átjutás, majd
a kérődzőkben megindult párhuzamos evolúció ideje már jobban
beazonosítható. Ennek viszonylag régen, de a kérődzők elkülönülése
után kellett bekövetkeznie, mert a többi emlősállatban nem
találhatók atadenovírusok, míg a kérődzőkben (szarvasmarhában,
juhban, kecskében, szarvasokban) felismert adenovírusoknak mintegy a
fele ide tartozik.
Megtörtént és várható változások
a hivatalos vírusrendszertanban
Kezdetben hihetetlennek tartott eredményeinket hosszas csatározások
és sok-sok év után sikerült átvinnünk a köztudatba, és hivatalosan
elfogadtatni a Nemzetközi Vírusrendszertani Bizottsággal (ICTV),
amelynek Adenoviridae Munkacsoportját 1999 óta intézetünk kutatói
vezetik. Munkánk alapján az Adenoviridae család eredeti két
nemzetsége mellett további három létesítését hagyta jóvá az ICTV.
Így a pillanatnyilag meglévő öt nemzetség a zárójelben említett
gazdákból származó vírusokat tartalmazza: Ichtadenovirus (egyetlen
hal-adenovírus), Siadenovirus (béka, Sulawesi-teknős és több madár),
Atadenovirus (pikkelyes hüllők, kérődzők, egy erszényes és madarak),
Aviadenovirus (csak madarak) és Mastadenovirus (csak emlősök)
(Harrach et al., 2011). Az egyes csoportok között a jelentős
eltérések a genom két végén elhelyezkedő, úgynevezett korai régiók
génjeiben vannak, míg a DNS középső részének szerveződése az egész
családra jellemző (Davison et al., 2003).
A Siadenovirus nemzetség eredete után kutatva
amerikai kollégákkal szinte azonos időben egy újabb vírusvonal első
képviselőit mutattuk ki ékszerteknősökben. A diagnosztikai PCR-ek
során alkalmazott primerek segítségével mindössze két, nagyon
megőrzött gén egy-egy rövid részletének szekvenciáját sikerült
meghatároznunk, de a filogenetikai számítások már ezek alapján is
egyértelműen mutatják, hogy a korábban ismert valamennyi
adenovírustól jelentősen eltérő, külön csoport tagjait ismertük fel.
Az ékszerteknősök mellett további, közeli rokon fajok egyedeiben
hasonló vírusokat mutattak ki mások is. A vizsgált DNS-szakaszok
A+T-tartalma kiegyensúlyozottnak bizonyult, és ennek alapján joggal
feltételezhető, hogy az új víruscsoport a (testudinoid) teknősökkel
együtt fejlődött leszármazási vonalnak felel meg. Annak
bizonyítására, hogy amint a törzsfa-rekonstrukciók sugallják, ezek a
vírusok valóban nemzetségszintű elkülönítést igényelnek, legalább
egy mintából a vírus teljes genomszekvenciájának meghatározása lenne
szükséges. Noha a vírus izolálására tett kísérleteink eddig
eredménytelenek voltak, jól tervezett PCR-ek segítségével talán
mégis sikerül majd a genom hosszabb részleteinek kinyerése is, és
így megismerhetjük majd a genomvégeknek feltehetően az új
nemzetségre specifikus tartalmát is.
Folyamatban lévő projektek,
nemzetközi együttműködések, további tervek
Jelenlegi munkahipotézisünk további tesztelése érdekében folytatjuk
széles körű felméréseinket, azaz valóságos „adenovírus-vadászatot”
valósítunk meg a vadon élő, minél eltérőbb evolúciós vonalakhoz
tartozó állatok körében. E célból számos ország kutatóival állunk
együttműködésben. Részt vettünk egy németországi denevér-adenovírus
izolátum jellemzésében és leírásában (Kohl et al., 2012), és hazai,
valamint német mintákban nagyszámú új adenovírus jelenlétét
bizonyítottuk. A denevér-adenovírusok kitűnő példát szolgáltatnak a
(gerinces osztályon belüli) lehetséges gazdaváltásokra. A vírusgenom
változékony régióinak összehasonlító elemzése alapján
megállapítottuk, hogy az egyik legrégebb óta ismert adenovírusos
betegség, nevezetesen a kutyák fertőző májgyulladásának vírusa és a
simaorrú denevérekben (Vespetilionidae) található adenovírusok igen
közeli közös őstől származnak. Feltételezzük, hogy a kutyákban
obligát patogénnek tekinthető, 1-es szerotípusú kutya-adenovírus nem
túl régen, már a modern időkben kerülhetett át denevérből a
ragadozókra (Carnivora), ezért nem látunk számottevő különbséget a
kutya és a különféle egyéb gazdák (farkas, róka, sakál, bűzös borz,
mosómedve, barna medve) adenovírusai között (Kohl et al., 2012).
Egyik doktoranduszom mostanában járt Brazíliában, hogy az Amazonas
vidékéről gyűjtsön különleges vadállatokból mintákat. Az OTKA
támogatásával közös kutatásokat indítunk a São Paulo-i Egyetemen
dolgozó kollégákkal a Dél-Amerikában honos gerincesek, így újvilági
majmok, vérszívó denevérek, vendégízületesek (hangyász, lajhár,
övesállat), kolibrik és más egzotikus madarak, hüllők stb.
adenovírusainak felderítésére.
Érdekes módon, noha elektronmikroszkópos
vizsgálatok alapján feltételezik krokodil-adenovírusok létezését is,
krokodilfélékből (Crocodilia) a mai napig nem sikerült
adenovírus-szekvenciát nyernünk, így ezek molekuláris jellemzése még
várat magára. Pedig ebben a madarakhoz igen közeli, gerinces
vonalban esetleg előforduló adenovírusok nagy érdeklődésre
tarthatnak számot. Elméletileg az sem zárható ki, hogy éppen a
siadenovírusok bizonyulnak majd e hüllőrenddel együtt evolválódott
vonalnak.
Potenciálisan a gyakorlatban is hasznosuló eredmények
Azt hiszem, kijelenthetjük, hogy az állati adenovírusok
összehasonlító vizsgálata és rendszertana napjainkban valódi
hungarikumnak tekinthető. Ezen a területen a magyar állatorvosok
úttörő szerepét és több évtizedes, folyamatos vezető pozícióját
világszerte ismerik és elismerik. Megtisztelő kötelességünknek
tartjuk e jó hírnév terjesztését és fenntartását. Erre már csak
azért is esélyünk van, mert mostanában, a még mindig romló gazdasági
viszonyok között, sajnos több olyan európai laboratóriumot
felszámoltak, ahol vadállatok kórokozóinak kutatása folyt.
Adenovírusok kimutatásának igényével folyamatosan keresnek meg
minket mind hazai, mind külföldi kollégák. Kiváltságosnak tartom
magam, hogy alkalmam van olyan, izgalmas kutatásokat folytatni,
amelyek számos szakdolgozónak, TDK-munkát végző diáknak és
doktorandusznak adtak és adnak lehetőséget az általuk legkedveltebb
állatcsoport virológiai vizsgálatára.
Ugyanakkor ezeknek a tiszta (sőt esetleg
haszontalan) alapkutatásnak tetsző vizsgálatoknak valójában
gyakorlati jelentőségük lehet. Az adenovírusok ugyanis általában
mind saját génjeiket, mind pedig a genomjukba mesterségesen
beültetett idegen géneket rendkívüli hatékonysággal képesek
kifejezni, és ezért a génterápiás eszközök és új típusú, rekombináns
vakcinák előállítása területén igen kedvelt, ha nem épp a
legkedveltebb modellnek számítanak. Állatorvosi vonalon többek
között a szarvasmarha, sertés, kutya és házityúk adenovírusaiból
próbáltak különféle rekombináns vakcinákat előállítani egyéb virális
kórokozók (például a veszettség vírusa) elleni immunizáláshoz.
Humán egészségügyi területen eleinte az emberi
adenovírusok vektorként történő alkalmazásának lehetőségeit
vizsgálták, de mivel a világ lakosságának nagy része természetes
fertőzések következtében az adenovírusok egynél több szerotípusa
ellen immunizálódik felnőtt korára, valós kockázat, hogy a majdan
kezelendő páciensekben meglévő, specifikus ellenanyagok
meghiúsíthatják a készítmények eredményes használatát. Ezért több
laboratórium és vállalat kezdett kísérletekbe olyan állati
adenovírusokkal, amelyekkel szembeni ellenanyagtól az emberi
populáció biztosan mentes. Ritka előfordulása és különleges
genomszerveződése miatt az Ausztráliában felfedezett, 7-es
szerotípusú juh-atadenovírust évek óta vizsgálják és manipulálják
különféle daganatsejtek felismerése és célzott eliminálása céljából.
A humán prosztatarák gyógyítására kifejlesztett, rekombináns
vírussal már klinikai teszteket is végeztek. A világon először
hazánkban Bartha Adorján által izolált atadenovírus, a 4-es típusú
szarvasmarha-adenovírus genomjának teljes nukleotidsorrendjét mi
határoztuk meg. A virion fíber nevű (antennaszerű) nyúlványának
génszakaszát francia együttműködésben humán gyógyászati célra szánt
(5-ös szerotípus) humán adenovírus genomjába építettük, hogy annak
tropizmusát megváltoztassuk (Renaut et al., 2004). Mivel a vírus
sejthez való elsődleges kapcsolódását a fíber közvetíti, e
fehérjének további módosításával (megfelelő ligandok
hozzákapcsolásával) tetszőlegesen választott sejtek specifikus
felismerésére alkalmas vírusvektor állítható elő.
További különleges állati adenovírusok (vagy ezek egyes génjeinek)
használhatóságát a humán gyógyászatban vakcinázás, génpótlás,
daganatos sejtek elpusztítása stb. céljából az elkövetkező években
európai uniós támogatásokkal kutathatjuk alaposabban olyan
kutatóhálózatokban és konzorciumokban, amelyekben a brit, finn,
francia, holland, spanyol, svájci és svéd résztvevők mellett
egyedüli állatorvosi és kelet-európai laboratóriumként veszünk
részt.
Támogatás: OTKA K72484, EU FP7-290002 ADVance Marie-Curie Initial
Training Network.
Kulcsszavak: adenovírus, evolúció, gazdaváltás, hüllők, kérődzők,
koevolúció, madarak
IRODALOM
Bartha Adorján (1969): Proposal for
Subgrouping of Bovine Adenoviruses. Acta Veterinaria Hungarica. 19,
319–321.
Benkő Mária (2008): Adenoviruses:
Pathogenesis. In: Mahy, Brian W. J. – van Regenmortel, Marc H. V.
(eds): Encyclopedia of Virology. Vol. 1. Third Edition. Elsevier,
Oxford, 24–29.
Benkő Mária – Harrach Balázs (1998): A
Proposal for a New (Third) Genus within the Adenoviridae Family.
Archives of Virology. 143, 829–837.
Benkő Mária – Bartha A. – Wadell, G.
(1988): DNA Restriction Enzyme Analysis of Bovine Adenoviruses.
Intervirology. 29, 346–350.
Benkő Mária – Élő P. – Ursu K. et al.
(2002): First Molecular Evidence for the Existence of Distinct Fish
and Snake Adenoviruses. Journal of Virology. 76, 10056–10059.
Benkő Mária – Harrach B. – D’Halluin, J.
C. (1990): Molecular Cloning and Physical Mapping of the DNA of
Bovine Adenovirus Serotype 4: Study of the DNA Homology among
Bovine, Human and Porcine Adenoviruses. Journal of General Virology.
71, 465–469.
Davison, Andrew J. – Benkő M. – Harrach B.
(2003): Genetic Content and Evolution of Adenoviruses. Journal of
General Virology. 84, 2895–2908.
Davison, Andrew J. – Wright, K. M. –
Harrach B. (2000): DNA Sequence of Frog Adenovirus. Journal of
General Virology. 81, 2431–2439.
Farkas Szilvia L. – Harrach B. – Benkő M.
(2008): Completion of the Genome Analysis of Snake Adenovirus Type
1, a Representative of the Reptilian Lineage within the Novel Genus
Atadenovirus. Virus Research. 132, 132–139.
Harrach Balázs (2000): Reptile
Adenoviruses in Cattle? Acta Veterinaria Hungarica. 48, 485–490.
Harrach Balázs – Benkő M. – Both, G. W. et
al. (2011): Family Adenoviridae. In: King, Andrew M. Q. – Adams, M.
J. – Carstens, E. B. – Lefkowitz, E. J. (eds.): Virus Taxonomy:
Classification and Nomenclature of Viruses. Ninth Report of the
International Committee on Taxonomy of Viruses. Elsevier, San Diego,
125–141.
Harrach Balázs – Meehan, B. M. – Benkő M.
et al. (1997): Close Phylogenetic Relationship between Egg Drop
Syndrome Virus, Bovine Adenovirus Serotype 7, and Ovine Adenovirus
Strain 287. Virology. 229, 302–306.
Ivanics Éva – Palya V. – Glávits R. et al.
(2001): The Role of Egg Drop Syndrome Virus in Acute Respiratory
Disease of Goslings. Avian Pathology. 30, 201–208.
Kohl, Claudia – Vidovszky M. Z. –
Mühldorfer, K. et al. (2012): Genome Analysis of Bat Adenovirus 2:
Indications of Interspecies Transmission. Journal of Virology. 86,
1888–1892.
Kovács M. Gábor – LaPatra, S. E. –
D’Halluin, J. C. – Benkő M. (2003): Phylogenetic Analysis of the
Hexon and Protease Genes of a Fish Adenovirus Isolated from White
Sturgeon (Acipenser transmontanus) Supports the Proposal for a New
Adenovirus Genus. Virus Research. 98, 27–34.
Papp Tibor – Fledelius, B. – Schmidt, V.
et al. (2009): PCR-Sequence Characterization of New Adenoviruses
Found in Reptiles and the First Successful Isolation of a Lizard
Adenovirus. Veterinary Microbiology. 134, 233–240.
Renaut, Laurence – Colin, M. – Leite, J.
P. et al. (2004): Abolition of hCAR-Dependent Cell Tropism Using
Fiber Knobs of Atadenovirus Serotypes. Virology. 321, 189–204.
Wellehan, James F. X. – Johnson, A. J. –
Harrach B. et al. (2004): Detection and Analysis of Six Lizard
Adenoviruses by Consensus Primer PCR Provides Further Evidence of a
Reptilian Origin for the Atadenoviruses. Journal of Virology. 78,
13366–13369.
LÁBJEGYZET
* Az MTA Bioinformatikai
Osztályközi Állandó Bizottság által, A bioinformatika Magyarországon
címmel szervezett tudományos ülésszakon, 2012. május 15-én,
Budapesten elhangzott előadás alapján készült szöveg.
<
|
|