|
jöttek létre (Bielz, 1861; Wagner, 1913; Soós,
1943; Szekeres, 2007). Tehát a vita tárgya tulajdonképpen az, hogy a
csavarodási irány homológ, avagy homoplasztikus bélyeg-e. Ez a
probléma azért maradhatott ennyi időn keresztül megoldatlan, mert e
kérdés eldöntésére a taxonómia klasszikus eszköztára nem volt
alkalmas.
Arra számítottunk, hogy egy viszonylag változékony
DNS-szakasz szekvenciaadatainak összehasonlítása révén eldönthető
lesz, hogy az azonos irányba csavarodó taxonok ténylegesen közeli
rokonai-e egymásnak, avagy sem. Emellett azt is szerettük volna
megbecsülni, hogy mikor keletkezhetett az Alopia génusz, mikor
váltak szét a főbb intragenerikus fejlődési ágak. Választásunk a
leggyakrabban vizsgált barcoding szakaszra, a mitokondriális
citokróm oxidáz génre esett, miután ez az a DNS-szakasz, amelyikről
az Alopia rokongénuszai tekintetében a legtöbb szekvenciaadat állt
rendelkezésre. Mintát vettünk az összes ismert alfaj reprezentatív
populációiból, és ezek alapján rekonstruáltuk a génusz molekuláris
törzsfáját. Az Alopia génusszal rokon sziklalakó orsócsigák
nagyrészt dél-európai elterjedésűek. Ezek génuszon belüli genetikai
variabilitása nagyobb mértékű, mint amit az Alopia esetében
találtunk. Ez arra utal, hogy az Alopia nemzetség a balkáni
rokonainál fiatalabb kialakulású, radiációjának kezdete valamikor a
korai vagy középső pleisztocén időszakra tehető. Eredményeink azt
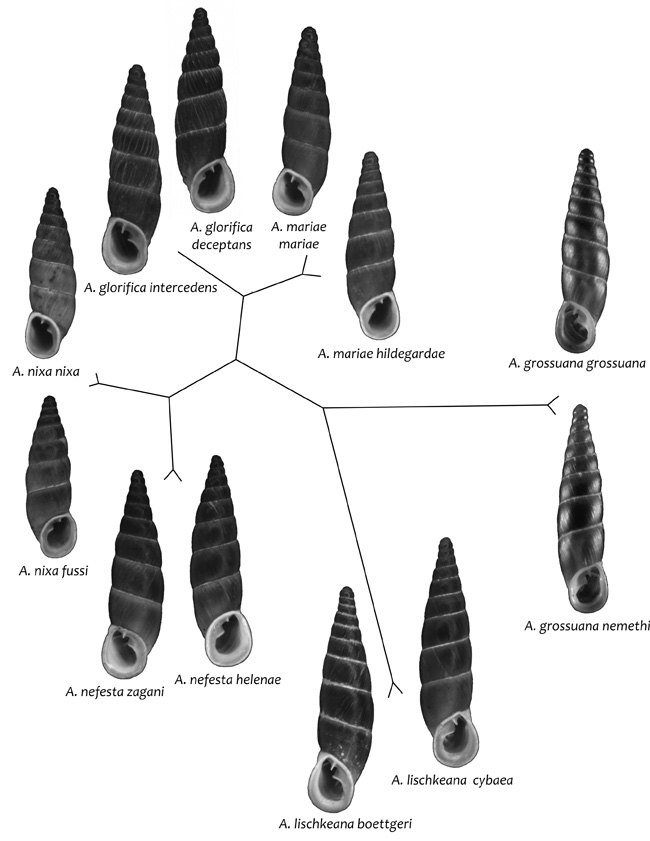
mutatták, hogy a jobbra, illetve balra csavarodó alfajok nem
alkotnak monofiletikus fejlődési vonalat
(1. ábra). A
DNS-szekvenciákból származtatott törzsfa alapján a génusz
törzsfejlődése során számos alkalommal, legalább tizenháromszor
fordult a kiralitás. Emiatt az európai csigáknál egyedülálló
tulajdonság miatt azt feltételezzük, hogy egy olyan csoportra
bukkantunk, amelynél a kiralitás genetikai determináltsága a
csigáknál eddig ismertekhez képest kevésbé stabil.
A törzsfa alapján az is nyilvánvaló, hogy a tükörképi taxonpárok
egymásnak igen közeli rokonai, miután a vizsgált szekvenciáiban ezek
közt nagyfokú, 96–100% egyezést találtunk. Ez azt jelzi, hogy a
tükörképi párok a fajkeletkezés korai fázisában lévő fiatal taxonok,
és egy filogenetikai alapon nyugvó rendszerben ezeket egy fajba
tartozó, külön alfajokként kell kezelnünk. Emiatt szükséges lesz a
génusz revíziója.
A fenti vizsgálatunk jó példa arra, miként
alkalmasak a molekuláris markerek alapján rekonstruált törzsfák
olyan problémák megoldására, amelyeknél a taxonómia klasszikus
módszerei önmagukban nem elégségesek. Jó szolgálatot tehetnek a
homológ és homoplasztikus morfológiai jegyek közti
különbségtételben, támpontként szolgálhatnak rokonsági kapcsolatok
feltárásánál, fajhatárok pontos kijelölésénél, valamint egyes
fejlődési ágak időbeni szétválásának meghatározásánál is.
Csodaszernek ugyanakkor a molekuláris filogenetikai analízis sem
tekinthető: mint bármely más módszernek, ennek is megvannak a maga
korlátai. Összességében azt mondhatjuk, hogy mára a molekuláris
markerek vizsgálata olyan fontos módszerré vált a taxonómia
eszköztárában, amely jól kiegészíti, de nem helyettesítheti a
tradicionális, morfológiai alapú vizsgálatokat a taxonómiában.
Kulcsszavak: puhatestű, csiga, Alopia, taxonómia, filogenetika,
DNS, citokróm oxidáz
IRODALOM
Adams, Dean C. – Rohlf, F. J. – Slice, D.
E. (2013): A Field Comes of Age: Geometric Morphometrics in the 21st
Century. Hystrix. 24, 7–14.
Alabadi, David – Blazquez, M. A. –
Carbonell, J. – Ferrandiz, C. – Perez-Amador, M. A. (2009):
Instructive Roles for Hormones in Plant Development. International
Journal of Developmental Biology. 53, 1597–1608.
Avise, John C. (2004): Molecular Markers,
Natural History, and Evolution (Second Edition). Sinauer,
Sunderland, MA
Bank, Ruud A. (2013): Fauna Europaea:
Gastropoda. Fauna Europaea version 2.6 •
WEBCÍM
Bielz, Eduard Albert (1861): Vorarbeiten
zu einer Fauna der Land- und Süsswasser-Mollusken Siebenbürgens.
Verhandlungen und Mitteilungen des Siebenbürgischen Vereins für
Naturwissenschaften zu Hermannstadt. 12, 83–179. •
WEBCÍM
Cheon, Jinyeong – Fujioka, S. – Dikles, B.
P. – Choe, S. (2013): Brassinosteroids Regulate Plant Growth through
Distinct Signaling Pathways in Selaginella and Arabidopsis. PLoS
ONE. 8, e81938.
Diverio, Silvana – Goddard, P. J. –
Gordon, I. J. (1996): Use of Long-acting Neuroleptics to Reduce the
Stress Response to Management Practices in Red Deer. Applied Animal
Behavior Science. 49, 83–88.
Fehér Zoltán – Németh L. – Nicoară, A. –
Szekeres M. (2013): Molecular Phylogeny of the Land Snail Genus
Alopia (Gastropoda: Clausiliidae) Reveals Multiple Inversions of
Chirality. Zoological Journal of the Linnean Society. 167, 259–272.
Grossu, Alexandru V. (1981): Gastropoda
Romaniae. Ordo Stylomatophora 3. Suprafamilile Clausiliacea şi
Achatinacea. Bucharest: Univrsitatea din Bucureşti.
Hebert, Paul D. N. – Cywinska, A. – Ball,
S. L. – deWaard, J. R. (2003): Biological Identifications through
DNA Barcodes. Proceedings of the Royal Society of London B. 270,
313–321. DOI: 10.1098/rspb.2002.2218
Kimakowicz, Mauritius von (1894):
Prodromus zu einer Monographie des Clausilia-Subgenus Alopia H. et
A. Adams. Verhandlungen und Mitteilungen des Siebenbürgischen
Vereins für Naturwissenschaften zu Hermannstadt, 43, 19–58.
Larsen, Thomas O. – Smedsgaard, J. –
Nielsen, K.F. – Hansen, M.E. – Frisvad, J.C. (2005): Phenotypic
Taxonomy and Metabolite Profiling in Microbial Drug Discovery.
Natural Product Reports. 22, 672–693. DOI: 10.1039/B404943H
Nordsieck, Hartmut (2008): The System of
the Genus Alopia H. & A. Adams 1855 (Gastropoda: Stylommatophora:
Clausiliidae). Mitteilungen der Deutschen Malakozoologischen
Gesellschaft. 79–80, 7–18.
Soós Lajos (1943): A Kárpát-medence
Mollusca-faunája (Mollusca Fauna of the Carpathian Basin).
Akadémiai, Budapest
Szekeres Miklós (2007): Four New
Subspecies of Alopia H. & A. Adams 1855 (Gastropoda, Pulmonata:
Clausiliidae). Schriften zur Malakozoologie. 23, 7–18.
Wagner, Antoni J. (1913): Die Familie der
Clausiliidae. In: Rossmässler, Emil Adolf: Rossmässler’s
Iconographie der Land- & Süsswasser-Mollusken, 21. Kreidel,
Wiesbaden •
WEBCÍM
Zimmermann, Niklaus E. – Edwards, T. C. –
Graham, C. G. – Pearman, P. B. – Svenning, J-C. (2010): New Trends
in Species Distribution Modelling. Ecography, 33, 985–989.
DOI: 10.1111/j.1600-0587.2010.06953.x
URL1
|