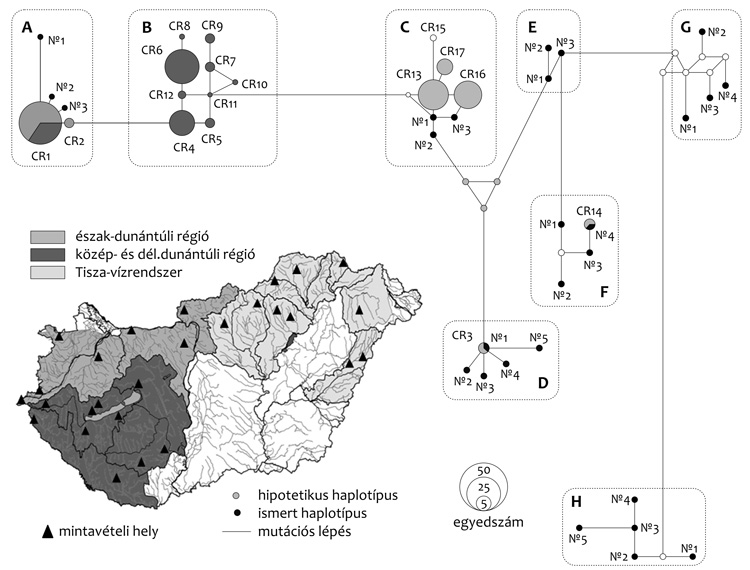
|
kerültek elő. Kisebb számban megjelentek a közép-dunántúli régió
vizeiben is. A közép- és dél-dunántúli régió vizeiben egy a dunai
küllőhöz hasonló, de annál jóval diverzebb haplotípuscsoport volt a
domináns („B” haplotípus csoport, 1. ábra). A két haplocsoport
elválását véleményünk szerint a pleisztocén „nagy
interglaciálisában” körülbelül 140 ezer éve bekövetkezett vízrajzi
szétkülönülés okozhatta, amikor a Közép- és Dél‑Dunántúl
vízrendszere izolálódott az észak-dunántúli vízrendszertől (Gábris –
Mari, 2007). Viszont a közép- és dél-dunántúli állományok nagyfokú
genetikai hasonlósága annak tudható be, hogy e két régió (ti. a
Dráva és a Balaton) vízrendszere csak jóval később, a holocénben
kezdett elkülönülni. Az a tény, hogy a CR01-es haplotípus a közép-
és dél-dunántúli területekről is előkerült, feltételezésünk szerint
másodlagos, emberi hatásoknak – véletlen betelepítéseknek – tudható
be.
Kevésbé változatos eredményeket hozott a Tisza vízrendszeréből
származó minták elemzése, amelyet véleményünk szerint a jóval
egyszerűbb felépítésű, és így könnyebben átjárható vízrendszerrel
magyarázhatunk. Összesen öt haplotípust tudtunk kimutatni, ezek
közül két, a Bózsvából előkerült egyed haplotípusa (CR14)
megegyezett a GenBankban EU131559-es kódszámmal szereplő Gobio
carpathicus haplotípussal. A többi tiszai vízgyűjtőről származó
minta a Jan Mendel és munkatársai (2008) által említett, de
önálló fajként le nem írt Gobio sp1 haplotípusaival mutattak nagy
hasonlóságot.
Az egymástól csak néhány mutációban különböző haplotípusokból
csoportokat képeztünk (ezeket az 1. ábrán bekereteztük és
betűkódokkal jelöltük). A csoportok elválását varianciaanalízissel
teszteltük (AMOVA) (Excoffier et al., 1992). Eredményeink azt
mutatják, hogy az összes haplocsoport szignifikánsan elkülönül
egymástól. Ugyanakkor a csoportok különbségei nem jelentősek. A
legnagyobb különbséget mutató G. obtusirostris („A” haplocsoport) és
G. insuyanus („H” haplocsoport, 3. táblázat) között is körülbelül
5%-nyi báziseltérés mutatható ki. Ilyen kismértékű genetikai
eltérések más fajok esetében (példéul: pénzes pér, kövi csík) csak
alfaji elkülönítést igazoltak (Marić et al., 2012; Šedivá et al.,
2008), így a kapott eredményeink megkérdőjelezik a közép-európai
küllőfélék (Gobio genus) faji szintű elkülönítésének jogosságát is.
Vizsgálataink eredményei rámutatnak, hogy a Kárpát-medence belső
területein élő pataklakó küllőfélék igen jelentős genetikai
sokféleséget mutatnak. Amellett, hogy három már leírt küllőfaj
haplotípusait jeleztük a területről (G. gobio, G. carpathicus, G.
obtusirostris), ezek mellett két elkülönülő rejtett taxonómiai
egységet is ki tudtunk mutatni a tiszai vízgyűjtőről, illetve a
közép- és dél-dunántúli régiókból, ahol ezek voltak a dominánsak.
Vizsgálatainkkal tehát egy ma is zajló természetes fajképződési
folyamat pillanatképét tudtuk rögzíteni, amelyet még kevéssé
befolyásoltak az emberi hatások (véletlen áttelepítések). A
konzervációbiológia és természetvédelem fő feladata, hogy ezt az
emberi léptékkel mérve igen hosszú idő alatt kialakult genetikai
mintázatot az élőhelyek védelmével óvja meg, illetve a szándékos
vagy véletlen áttelepítések megakadályozásával fenntartsa ezeknek az
így kimutatott rejtett (kriptikus) taxonómiai egységeknek a
genetikai izolációját.
Kulcsszavak: kriptikus fajok, molekuláris taxonómia,
konzervációbiológia, mitokondriális haplotípusok
IRODALOM
Bănărescu, Petre M. – Sorić, V. M. –
Economidis, P. S. (1999): Gobio gobio (Linnaeus, 1758). In:
Bănărescu, Petre M. (ed.): The Freshwater Fishes of Europe. Vol.
5/I. Cyprinidae 2. Part 1. Rhodeus to Capoeta. AULA, Wiebelsheim,
81–134.
Bandelt, Hans-Jürgen – Forster, P. – Röhl,
A. (1999): Median-joining Networks for Inferring Intraspecific
Phylogenies. Molecular Biology and Evolution 16, 37–48. •
WEBCÍM
Dudgeon, David – Arthington, A. H. –
Gessner, M. O. – Kawabata, Z. I. – Knowler, D. J. et al., (2006):
Freshwater Biodiversity: Importance, Threats, Status and
Conservation Challenges. Biological Reviews. 81, 163–182.
DOI: 10.1017/S1464793105006950
Erős Tibor (2007): Partitioning the
Diversity of Riverine Fish: The Roles of Habitat Types and
Non-native Species. Freshwater Biology. 52, 1400–1415. DOI: 10.1111/
j.1365-2427.2007.01777.x
Excoffier, Laurent – Smouse, E. P. –
Quattro, M. J. (1992): Analysis of Molecular Variance Inferred from
Metric Distance among DNA Haplotypes: Application to Human
Mitochondrial DNA Restriction Data. Genetics. 131, 479–491. •
WEBCÍM
Gábris Gyula – Mari László (2007): A
Zala-folyó lefejezése. Földrajzi Értesítő. 56, 1–2, 39–50. •
WEBCÍM
Kottelat, Maurice – Freyhof, Jörg (2007):
Handbook of European Freshwater Fishes. Publications Kottelat,
Cornol, Switzerland
Marić, Saša – Kalamujić, B. – Snoj, A. –
Bajec, S. S. (2012): Genetic Variation of European Grayling
(Thymallus thymallus) Populations in the Western Balkans.
Hydrobiologia. 691, 225–37. DOI: 10.1007/s10750- 012-1076-2 •
WEBCÍM
Mayr, Ernst (1948): The Bearing of the New
Systematics on Genetical Problems. The Nature of Species. Advances
in Genetics. 2, 205–237
Mendel, Jan – Lusk, S. – Vasil’eva, E. D.
– Reshetnikov, S. I. (2008): Molecular Phylogeny of the Genus Gobio
Cuvier, 1816 (Teleostei: Cyprinidae): and Its Contribution to
Taxonomy. Molecular Phylogenetics and Evolution. 47, 1061–1075. DOI:
10.1016/j.ympev.2008.03.005
Šedivá, Alina – Janko, K. – Šlechtová, V.
– Vassilev, M. (2008): Around or across the Carpathians:
Colonization Model of the Danube Basin Inferred from Genetic
Diversification of Stone Loach (Barbatula barbatula): Populations.
Molecular Ecology. 17, 1277–1292.
DOI: 10.1111/j.1365-294X.2007.03656.x
Takács Péter (2012): Morphometric
Differentiation of Gudgeon Species Inhabiting the Carpathian Basin.
Annales de Limnologie – International Journal of Limnology. 48,
53–61. DOI: 10.1051/limn/2011058 •
WEBCÍM
URL1
|