|
|
Bevezetés
A nappali lepkék a legnépszerűbb élőlénycsoportok közé tartoznak.
Ennek is köszönhető, hogy már a XX. század első felében is több
genus monografikus feldolgozása megjelent. Ezek közé tartozott
Lionel G. Higgins (1941) Melitaea-katalógusa, amely színes táblákon
örökítette meg az addig ismert palearktikus fajokat. Szintén Higgins
volt az első, aki ezen belül a phoebe-fajcsoportot definiálta.
Genitália- és szárnymorfológiai jellegek alapján hét fajt sorolt
ebbe a csoportba. Közülük legismertebb faj a Melitaea phoebe (Denis
& Schiffermüller, 1775), amelynek elterjedési területe
Észak-Afrikától egészen Kelet-Ázsiáig húzódik. A M. sibina
Alphéraky, 1881-et Közép- és Belső-Ázsia magashegységeiből ismerjük.
A csoport legnagyobb méretű tagja a M. scotosia Butler, 1878, amely
kizárólag a Távol-Keleten (Amur-vidék, Mandzsúria, Észak-Kína,
Korea, Japán) fordul elő. A M. aetherie (Hübner, 1826) a
Mediterráneum nyugati részének lakója, Szicíliában, az
Ibériai-félsziget déli részén és Észak-Afrika nyugati részein él.
Végül ide tartozik még három viszonylag szűk elterjedésű faj: a
közel-keleti M. turkmanica Higgins, 1940, M. consulis Wiltshire,
1941 és M. collina Lederer, 1836.
Az elmúlt évtizedek kutatásai sokat változtattak a
Higgins által fölvázolt képen. Kovács Lajos már az 1950-es években
rájött arra, hogy Magyarországon két eltérő phoebe alak található.
Ekkor még az átmeneti fenotípusú egyedek miatt csak alfaji szintű
elkülönítésre gondoltak (lásd: Melitaea phoebe kovacsi Varga, 1967).
A továbbiakban két kutatócsoport egymástól függetlenül ismerte fel a
két, átfedő elterjedésű faj életciklusbeli és hernyó-morfológiai
különbségeit. Kitűnt, hogy míg a M. phoebe elterjedési területének
klimatikus sajátságai alapján évente egy, illetve két nemzedéket hoz
létre (Közép- és Dél-Európában rendszerint kettőt), addig a
„rejtett” faj mindenütt, a Mediterráneumtól a Kárpát-medencéig,
egynemzedékű. Emellett olasz, görög, izraeli és magyarországi
anyagok vizsgálata arra az egybehangzó eredményre vezetett, hogy míg
a M. phoebe fejlett hernyójának fejtokja fekete, addig a felismert
„rejtett faj” fejlett hernyójának fejtokja vörös. Ennek
megjelölésére típusanyagok vizsgálata alapján Peter Russel és
munkatársai a M. telona Fruhstorfer, 1908 nevet (típuslelőhely:
Jeruzsálem) javasolták.
Újabb eredmények: molekuláris filogenetika
és geometriai morfometria
Az utóbbi évtized molekuláris filogenetikai vizsgálatai (Leneveu et
al., 2009) megmutatták, hogy a M. phoebe fajcsoport a Didymaeformia
alnemen belül monofiletikus egységet képez. Ez a vizsgálat
megerősítette, hogy a M. telona valóban önálló, a M. phoebe-től
eltérő faj. Emellett az is tisztázódott, hogy a korábban a M. phoebe
alfajának tekintett, sőt a M. telonával is összekevert M. punica
Oberthür, 1876 is önálló fajt képvisel. Egy további új közlemény
(Tóth – Varga, 2011) az ivarszervek geometriai („landmark”-alapú)
morfometriai vizsgálata alapján tovább finomította a M. phoebe
fajcsoportról alkotott képet. Azt is tisztázta, hogy a M. phoebe
eltérő alakjaként leírt, majd később alfajának tekintett M. phoebe
ornata Christoph, 1893 valójában a M. telona formakörbe tartozik, és
a nevezéktan prioritási szabálya alapján ez a szóban forgó faj
érvényes neve (Melitaea ornata Christoph, 1893 = M. telona
Fruhstorfer, 1908). Ez a közlemény mutatta meg, hogy a Melitaea
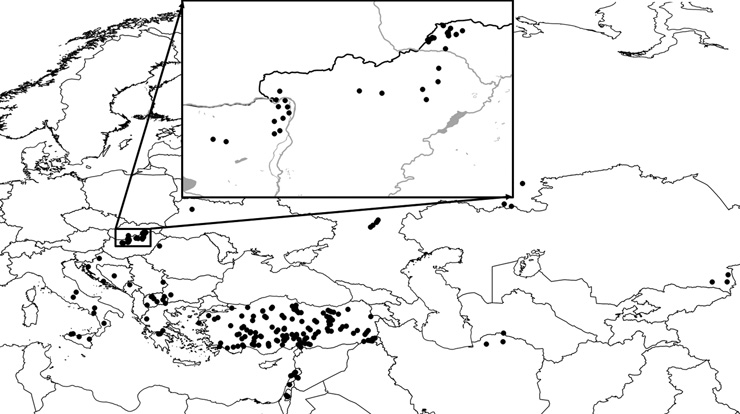
ornata elterjedése (1. ábra)
jóval nagyobb, mint azt előzőleg gondoltuk, mivel számos helyről
került elő Dél-Oroszországból és Kazahsztánból. Egyben világossá
vált, hogy a pontomediterrán-turkesztáni faj legészakibb
populációcsoportja a korábban a M. phoebe alfajaként leírt M. ornata
kovacsi, amely az erős földrajzi izoláción kívül
tápnövény-specializációja (Cirsium pannonicum) révén is eltér a faj
további populációitól.
A genitália morfometria eredményei nagyrészt
összhangban vannak a DNS-alapú vizsgálatokkal. Megerősítik a M.
punica faji státuszát, és azt mutatják, hogy a M. sibina mindössze a
M. phoebe egy jellegzetes magashegységi alakja. A távol-keleti M.
scotosia esetében viszont – a DNS-vizsgálatoknak ellentmondóan –
megerősítik annak
|
|
|
faji státuszát. A morfometriai analízisek egy
további eredménye egy eddig ismeretlen új faj leírása Irán
területéről (M. zagrosi Tóth & Varga, 2011), amely jelentős eltérést
mutat az eddig ismert fajoktól a szárnymintázatbeli különbségeken
kívül a hím és nőstény genitáliajellegek alapján egyaránt.
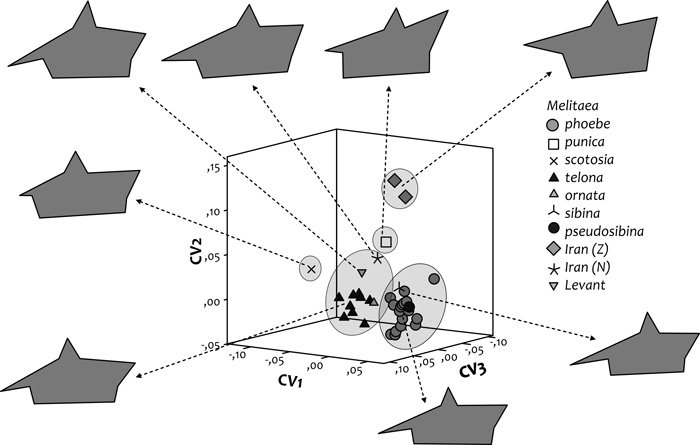
A morfometriai vizsgálatok alapján ordinációt
végeztünk, amelynek főbb eredményeit a
2. ábrán mutatjuk be. A diszkriminancia-analízis scatterplotján
a vizsgált taxonok kilenc landmarkpont alapján megszerkesztett
átlagos processus posterior alakjai láthatók. A morfometriai
vizsgálat azt is megmutatta (lásd Tóth – Varga, 2011), hogy a M.
ornata elterjedési területén belül négy fő morfotípusra tagolódik.
(i) A „nyugati” morfotípus a dél-itáliai,
Kárpát-medencei és délnyugat-ukrajnai populációkat foglalja magába.
(ii) A „keleti” csoport a legnagyobb elterjedésű: áreája a
Balkán-félsziget keleti részétől Kis-Ázsián és Dél-Oroszországon át
Kazahsztánig tart. (iii) A legnagyobb morfológiai távolságot mutató
levantei csoport (= M. ornata telona s. str.) aránylag szűk
területen, Libanon és Izrael területén él. (iv) Az észak-iráni
csoport szintén korlátozott elterjedésű, az Elbursz- és
Kopet-dagh-hegységek területéről ismerjük. A M. ornata elterjedési
modelljének felhasználásával predikáltuk a potenciális refúgiumokat,
ahol e faj a legutóbbi jégkorszak lehűlési időszakát vészelhette át.
A morfometriai eredmények és a potenciális refúgiumok összevetéséből
úgy tűnik, hogy a Kárpát-medence egy Appennini refúgiumból népesült
be (lásd Tóth et al., 2013). Ennek részleteit és a fajcsoport eddig
kevéssé vizsgált filogenetikai viszonyait azok a molekuláris
vizsgálatok hivatottak tisztázni, amelyeket a közelmúltban
indítottunk el.
Kitekintés
Úgy véljük, eredményeink rámutatnak arra, hogy a korszerű
módszerekkel végzett vizsgálatok lehetővé teszik a korábban, külső
morfológiai jellegek alapján rejtettnek látszó taxonómiai eltérések,
és ezzel a biológiai sokféleség egy finomabb szintjének
felismerését. Kitűnt, hogy a vizsgált fajcsoportban a
Kárpát-medencében egy sajátos életmódú, földrajzilag elkülönült
populációcsoport (alfaj) él, amelynek megőrzése természetvédelmi
intézkedéseket igényel.
Kulcsszavak: rejtett fajok, monofiletikus fajcsoport,
genitália-morfológia, molekuláris filogenetika, elterjedés,
életciklus
IRODALOM
Higgins, Lionel G. (1941): An Illustrated
Catalogue of the Palearctic Melitaea (Lep. Rhopalocera).
Transactions of the Royal Entomological Society of London. 91,
175–365. DOI: 10.1111/j.1365-3113.1944.tb00780.x
Leneveu, Julien– Chichvarkhin, A. –
Wahlberg, N. (2009): Varying Rates of Diversification in the Genus
Melitaea (Lepidoptera: Nymphalidae) during the Past 20 Million
Years. Biological Journal of the Linnean Society. 97, 346–361.
DOI: 10.1111/j.1095- 8312.2009.01208.x
Pecsenye Katalin – Bereczki J. – Tihanyi
B. et al. (2007): Genetic Differentiation among the Maculinea
species (Lepidoptera: Lycaenidae) in Eastern Central Europe.
Biological Journal of the Linnean Society. 91, 11–21.
DOI: 10.1111/j.1095-8312.2007.00781.x
Russell, Peter – Tennent, W. J. – Pateman,
J. – Varga Z. et al. (2007): Further Investigations into Melitaea
telona Frushstorfer, 1908 (=ogygia Frushstorfer, 1908 =emipunica
Verity, 1919) (Lepidoptera: Nymphalidae), with Observations on
Biology and Distribution. Entomologist’s Gazette. 58, 137–166.
Tóth János P. – Varga Zoltán (2010):
Morphometric Study on the Genitalia of Sibling Species Melitaea
phoebe and M. telona (Lepidoptera: Nymphalidae). Acta Zoologica
Academiae Scientiarum Hungaricae. 56, 273–282. •
WEBCÍM
Tóth János P. – Varga Zoltán (2011):
Inter- and Intraspecific Variation in the Genitalia of the Melitaea
phoebe Group (Lepidoptera, Nymphalidae). Zoologischer Anzeiger - A
Journal of Comparative Zoology. 250, 258–268. DOI:
10.1016/j.jcz.2011.05.002
Tóth János P. – Varga K. – Végvári Zs. –
Varga Z. (2011): Distribution of the Eastern Knapweed Fritillary
(Melitaea ornata Christoph, 1893) (Lepidoptera: Nymphalidae): Past,
Present and Future. Journal of Insect Conservation. 17, 245–255.
DOI: 10.1007/s10841-012-9503-2
|
|