|
határvidéken. A T. cristatus és T. dobrogicus fajok
elkülönítése a morfológia alapján problémás, amit még az is nehezít,
hogy az elterjedésük határán keverednek egymással. Kutatásunkban
molekuláris taxonómiai módszereket alkalmazva szerettük volna
pontosítani a három faj előfordulását Magyarországon, különös
tekintettel a T. cristatus – T. dobrogicus fajokra.
Összesen 140 tarajosgőte-példányt mintáztunk
Magyarország területéről (25 T. carnifex és 115 T. dobrogicus/T.
cristatus), majd a mitokondriális ND2-szakaszt és hét
mikroszatellita lókuszt vizsgáltunk a laboratóriumban. Eredményeink
azt mutatták, hogy hazánk legnagyobb területén a T. dobrogicus faj
fordul elő, míg a T. cristatus csak cristatus/dobrogicus-hibrid
példányok formájában van jelen az Aggteleki-karszt területén. A
nyugati peremvidékről származó T. carnifex populációk élesen
elkülönültek a többi állománytól a filogenetikai fán (Vörös – Major,
2007).
Munkánk következő lépcsőjében negyven fehérje
lókuszt vizsgáltunk a T. dobrogicus fajon, hiszen ennek a fajnak az
elterjedéstörténete még nem volt pontosan ismert. Összesen 140
egyedet mintáztunk 16 populációból, a Bécsi-medencétől a Duna alsó
szakaszáig. A populációgenetikai vizsgálatok magas
allélváltozatosságot és jelentős génáramlást mutattak a három
nagyobb folyó (Duna, Tiszta, Száva) mentén, amelyekhez a faj
előfordulása kötődik. A legmagasabb genetikai diverzitást a
Száva-menti populációk mutatták, amely arra utal, hogy ezen a
területen lehetett a faj egyik menedékterülete az utolsó eljegesedés
során (Vörös – Arntzen, 2010).
A barna varangy (Bufo bufo) filogeográfiája
Néhány évvel ezelőttig a Bufo volt az egyik legfajgazdagabb kétéltű
génusz, amely 283 ismert békafajt számlált. Darrell L. Frost és
munkatársai (2006) azonban csonttani, morfológiai és genetikai
vizsgálatokon alapulva több génuszra darabolták szét. A Bufo
génuszban maradt keleti és nyugati fajok is jelentős genetikai
különbséget mutattak. Vizsgálatunkban a molekuláris taxonómia
módszereivel kutattuk a Bufo bufo fajcsoport nyugati tagjainak (Bufo
bufo, Bufo eichwaldi, B. verrucosissimus) leszármazási viszonyait. A
három faj közül az eurázsiai elterjedésű Bufo bufo hazánkban is
gyakori a sík- és dombvidékeken egyaránt. A Bufo bufo fajnak eddig
három alfaját írták le: B. b. bufo, B. spinosus és B. b.
gredosicola. Vizsgálatunkban a B. bufo törzsalak és a B. spinosus
alfaj leszármazási viszonyaira és kontaktzónájára
összpontosítottunk.
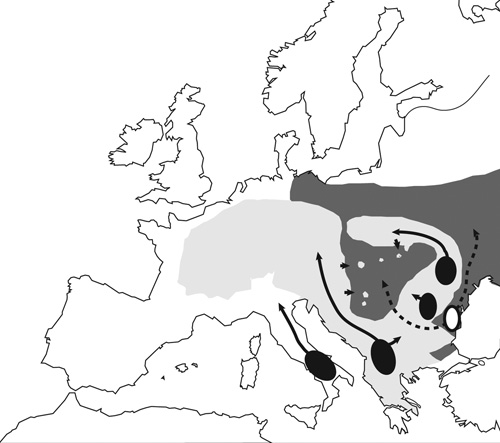
A három faj összesen 232 egyedét mintáztuk, lefedve
Európát, Észak-Afrikát, de gyűjtöttünk szövetmintát Anatóliától
egészen a Kaukázusig. Két mitokondriális génszakaszt (16S és
citokróm-b) és négy nukleáris gént (POMC, CXCR4, BDNF, RPL3)
szekvenáltunk. Külcsoportként egy közelrokon, a keleti fajcsoportból
származó faj (B. gargarizans) szekvenciáját használtuk fel.
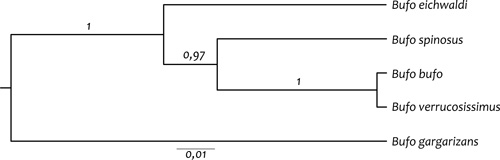
A filogenetikai rekonstrukció azt mutatta, hogy a
B. eichwaldi testvértaxonja egy kládusznak, amely tartalmazza a B.
b. spinosus alfajt, és egy másik kládusznak, amely tartalmazza a B.
b. bufo törzsalak és a közelrokon B. verrucosissimus mintákat (2.
ábra). Ezek alapján a törzsalak B. b. bufo és az
Észak-Afrikában, az Ibériai-félszigeten és Franciaország nyugati
felén előforduló B. b. spinosus génállományukban olyan jelentősen
különböznek, hogy önálló faji rangra érdemesek. A filogeográfiai
rekonstrukció és a molekuláris óra alapján a B. bufo és B. spinosus
kláduszokat nagyjából kilencmillió évvel ezelőtt, a korai Pireneusok
kiemelkedése izolálta el egymástól, amely időszaktól számítható a
két faj önálló evolúciós története (Recuero et al., 2012).
A Kárpát-medence további kétéltűfajainak filogeográfiai feldolgozása
folyamatban van. Jelenlegi kutatásaink a foltos szalamandra
(Salamandra salamandra) és az alpesi gőte (Ichthyosaura alpestris)
molekuláris taxonómiáját és elterjedéstörténetét vizsgálják.
Kulcsszavak: filogeográfia, mitokondriális DNS, nukleáris DNS,
mikroszatellitek, sárgahasú unka, vöröshasú unka, alpesi
tarajosgőte, dunai tarajos-gőte, közönséges tarajosgőte, barna
varangy
IRODALOM
Babik, Wieslav – Branicki, W. – Crnobrnja-
ISAILOVIĆ, J. et al. (2005): Phylogeography of Two European Newt
Species – Discordance between mtDNA and Morphology. Molecular
Ecology. 14, 2475–2491. DOI: 10.1111/j.1365-294X.2005.02605.x
Babik, Wieslav – Branicki, W. – Sandera,
M. et al. (2004): Mitochondrial Phylogeography of the Moor Frog,
Rana arvalis. Molecular Ecology. 13, 1469–1480.
DOI: 10.1111/j.1365-294X.2004.02157.x
Bucci-Innocenti, Stefania – Ragghianti, M.
– Mancino, G. (1983): Investigations of Kariology and Hybrids in
Triturus boscai and T. vittatus, with a Reinterpretation of the
Species Groups within Triturus (Caudata: Salamandridae). Copeia. 3,
585–598.
Dely Olivér Gy. (1967): Kétéltűek –
Amphibia – Magyarország Állatvilága (Fauna Hungariae), 20, 3.
Akadémiai, Budapest
Frost, Darrell R. – Grant, T. – Faivovich,
J. et al. (2006): The Amphibian Tree of Life. Bulletin of the
American Museum of Natural History. 297, 1–370. •
WEBCÍM
Pabijan, Maciej – Wandicz, A. – Hofman, S.
et al. (2013): Complete Mitochondrial Genomes Resolve the
Phylogenetic Relationships within Bombina (Anura: Bombinatoridae).
Molecular Phylogenetics and Evolution. 69, 63–74. DOI:
10.1016/j.ympev.2013. 05.007 •
WEBCÍM
Recuero, Ernesto – Canestrelli, D. – Vörös
J.et al. (2012): Multilocus Species Tree Analyses Resolve the
Radiation of the Widespread Bufo Bufo Species Group (Anura:
Bufonidae). Molecular Phylogenetics and Evolution. 62, 71–86. DOI:
10.1016/j.ympev.2011.09.008
Schmitt, Thomas – Varga Zoltán (2012):
Extra-Mediterranean Refugia: The Rule and Not the Exception?
Frontiers in Zoology. 9, 22. doi:10.1186/1742-9994-9-22 •
WEBCÍM
Vörös Judit – Alcobendas, M. –
Martínez-Solano, I. et al. (2006): Evolution of Bombina bombina and
Bombina variegata (Anura: Discoglossidae) in the Carpathian Basin: A
History of Repeated mt-DNA Introgression across Species. Molecular
Phylogenetics and Evolution. 38, 705–718. DOI:
10.1016/j.ympev.2005.08.010
Vörös Judit – Arntzen, Jan W. (2010): Weak Population Structuring in
the Danube Crested Newt, Triturus dobrogicus, Inferred from
Allozymes. Amphibia-Reptilia. 31, 339–346. •
WEBCÍM
Vörös Judit – Major Ágnes (2007):
Kétéltű-populációk földrajzi szerkezete a Kárpát-medencében. In:
Forró László (szerk.): A Kárpát-medence állatvilágának kialakulása.
Magyar Természettudományi Múzeum, Budapest
Wielstra, Ben – Litvinchuk, S. N. –
Naumov, B. et al. (2013): A Revised Taxonomy of Crested Newts in the
Triturus karelinii Group (Amphibia: Caudata: Salamandridae), with
the Description of a New Species. Zootaxa. 3682, 441–453. •
WEBCÍM
|