|
szelekció volt a nagyobb nemesítő intézetekben, így
Martonvásáron is. A modern nemesítési módszerek közül napjainkban az
új genetikai források létrehozására a molekuláris markerszelekció
széles körű alkalmazásának vagyunk szemtanúi.
A nemesítőknek alapvetően öt különböző genetikai
forrás áll rendelkezésükre az előnemesítésben, úgy, mint egy adott
régióban adaptív fajta, más ökológiai régióban termesztett ún.
exotikus genotípus, a génbankokban fellelhető régi tájfajta vagy
fajtapopuláció, vadon élő vagy termesztett rokon fajok, valamint a
mutáns genotípusok.
Adaptív fajták és törzsek felhasználásával a
legkisebb az esély a genetikai variabilitás jelentősebb
szélesítésére. Ez leginkább a vadon élő vagy termesztett rokon fajok
felhasználásával valósítható meg. Ugyanakkor ez utóbbi a hagyományos
genetikai módszerekkel időigényes kutatás. Elég, ha csak a
legnagyobb területen elterjedt 1B/1R rozs transzlokációs
búzafajtákat említjük példaként: az első sikeres búza×rozs
keresztezéstől az első fajta regisztrációjáig harminchárom év telt
el (Rabinovich, 1998). A nemesítési folyamat bonyolultsága ellenére
a búzanemesítők nagymértékben használtak fel vad fajokat.
Interspecifikus keresztezésekkel világszerte több
betegségrezisztencia- és tartalékfehérje-gént sikerült beépíteni a
közönséges búza genomjába.
Ökológiai régiónkban a búza egyik legfontosabb
tulajdonsága az átlagon felüli abiotikus stresszrezisztenciája. Ez
több komponensből tevődik össze, és ide tartozik a télállóság. A
martonvásári fitotronban végzett fagyállósági kísérletek során
vizsgálatba állítottuk az Európa különböző régiójából származó, az
adott régióra jellemző búzafajtákat (Veisz et al., 1996). A
kísérleti eredmények alapján a kelet-európai fajták bizonyultak a
legelenállóbbaknak, és a dél-európaiak a leginkább fagyérzékenyek. A
klímaváltozással még inkább felértékelődtek a szárazság- és a
hőtűréskutatások. Aszálynak kitett kontinentális klímájú régiókban
az adaptációs képesség, a szárazságtűrés és a termőképesség szorosan
összefüggő tulajdonságok. Különösen jelentőssé vált ez a probléma a
szélsőséges klimatikus események gyakoriságának fokozódásával, ami
nemcsak a termés mennyiségének, hanem minőségének stabilitását is
megkérdőjelezi. Ezáltal nemcsak az ökológiai régiók, de az évjáratok
közti fluktuáció is megnőtt.
A termésstabilitás másik fontos meghatározója a
gombabetegségekkel szembeni rezisztencia számos kórokozónál főként
mono- vagy oligogénes rezisztenciaöröklődésen alapul. Ez kiváló
lehetőséget nyújt a molekuláris marker technológia felhasználására a
rezisztencianemesítésben. A legfontosabb kórokozókkal (levélrozsda,
lisztharmat, sárgarozsda, szárrozsda stb.) szemben ellenálló
génforrások kimutatásával pontosan meg tudjuk határozni fajtáink,
fejlett törzseink és génforrásaink rezisztenciagénjeit. A hatékony
rezisztenciagének gyors beépítése érdekében az ún. molekuláris
marker segítségével végzett visszakeresztezéses nemesítést
alkalmazunk (marker-assisted back crossbreeding – MABC).
Felgyorsítható az azonos fenotípusos hatást előidéző allélok
piramidálása egy genotípusba, általában egy jól bevált búzafajtába,
amit egyetlen tulajdonságban kívánunk módosítani. A génpiramidálás
és a molekuláris markerek felhasználásával végzett
visszakeresztezéses nemesítés optimálisan kombinálható a hagyományos
szelekciós módszerekkel. Jó példa erre a BIOEXPLOIT FP6 EU-projekt
keretében végzett levélrozsda rezisztenciagének piramidálása
martonvásári búzafajtákba (Vida et al., 2009). Egy levélrozsdára
erősen fogékony, de a sikérmennyiségre és -minőségre egyaránt kiváló
fajtánkat, az Mv Emmát használtuk recipiens szülőként a program
során.
A búzafajták döntő többsége a világszerte elterjedt
észak-amerikai osztályozás alapján a piros, keményszemű (hard red)
minőségcsoportba tartozik. Az elmúlt két évtized során a csökkent
műtrágya-felhasználás, a szélsőséges klíma és a biotikus
stressztényezők hatásai miatt romlott a beltartalmi minőségi
paraméterek stabilitása. A kontinentális klimatikus éghajlati
környezetben nemesített búza minőségstabilitása azért is kritikus
tulajdonság, mivel ebben a régióban általában jobb sütőipari
minőségű, nagyobb fehérjetartalmú búzát lehet termeszteni, mint
például Nyugat-Európában. A martonvásári nemesítési program jelentős
eredményeket ért el ezen a téren. A nagy sikértartalmú búzafajták
nemesítése során sikerült kisebb nitrogénműtrágya-dózis
felhasználása esetén is megfelelő sikértartalmú termést produkálni
az Mv Magdaléna, az Mv Csárdás, az Mv Verbunkos fajtákkal. A
2002-ben állami elismerésben részesült Mv Suba mind
sikértartalomban, mind -minőségben Európa egyik legjobb búzafajtája,
amit tíz országban termelnek minőségjavítás céljából. Az új
nemesítési kiindulási anyag szelekciója során mindenképpen célszerű
figyelembe venni a kanadai növénynemesítők koncepcióját, miszerint a
fehérjetartalom további jelentős növelése helyett inkább a
fehérjekomponensek összetételének változtatása, minőségének javítása
jelenthet új törekvést (DePauw et al., 1998). Ebből kiindulva
született meg az Mv Kolo, az Mv Toldi, majd a Bánkúti 1201-es régi
magyar búzából átvitt Bx7 fehérjetúltermelő gént tartalmazó Mv
Karizma.
Az egészséges gabonaalapú táplálkozáshoz a búzaszem
bioaktív komponenseinek javítása is előtérbe került. A növényi
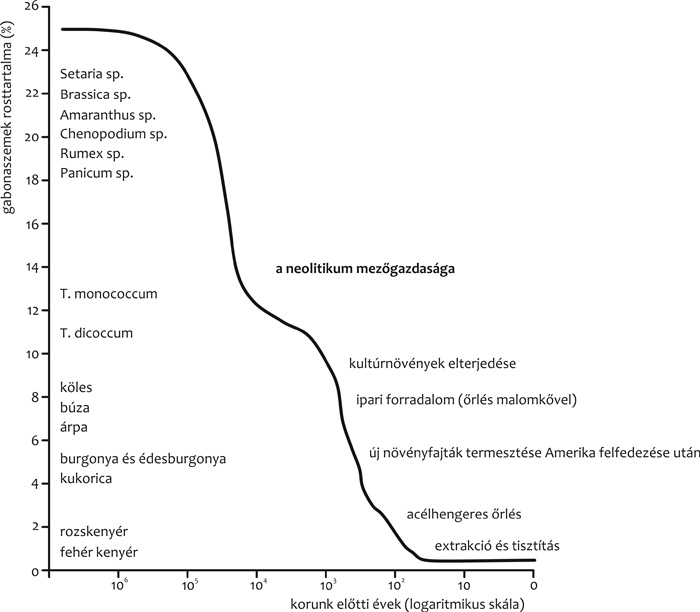
rostanyagok a modern gabonaalapú táplálkozásból sajnos a korszerű
malomipari technológiák miatt szorultak ki az emberiség történelme
során (1. ábra). A
klasszikus értelemben vett növényi rostanyagok – az arabinoxilán és
a β-glükán – a gabonaalapú élelmiszerek táplálkozástani értékét
nagymértékben befolyásolják, a búzaszemben az endospermium
sejtfalában találhatók meg. Így többek között szerepük van az
inzulinszabályozásban, csökkentik a koleszterinszintet, a 2-es
típusú diabétesz kialakulásának kockázatát. Ezidáig csak exotikus
forrásokban sikerült nagy arabinoxilán és a gyors felszívódás miatt
értékes vízoldható arabinoxilán- (WE-AX-) tartalmat kimutatni.
Közösen a rothamstedi és a Budapesti Műszaki Egyetem kutatóival
előállítottunk nagy WE-AX-tartalmú genotípusokat.
Egy másik jelentős, és eddig hazánkban kevésbé
vizsgált beltartalmi komponens, a búzaszem meghatározó összetevője a
keményítő, amit szintén a rostanyagok közé sorolnak. A
búzakeményítőben az amilóz és az amilopektin aránya megközelítőleg
1:3, a szerkezetbeli különbségek az amilóz és az amilopektin között
jelentősek, ami többek között befolyásolja a komponensek
emészthetőségét. A lassúbb emészthetőség humán egészségügyi
szempontból fontos tulajdonság, de emellett a búzakeményítőnek
számos más felhasználási lehetősége van, mint például a bioetanol
előállítása, az állati takarmányozás, műanyagok, filmek és
ragasztóanyagok előállítása stb. A HEALTHGRAIN FP6 EU-projekt
keretében sikerült mindkét irányba módosított keményítő összetételű
genotípusokat szelektálni, azaz kis amilóztartalmú, ún. waxy,
részleges waxy, valamint nagy, közel 40% amilóztartalmú genetikai
forrásokat előállítani molekuláris markerek felhasználásával
(Rakszegi et al., 2014).
A mutációs nemesítés „reneszánszát” is a
molekuláris technológiák elterjedése segítheti elő. A TILLING
(Targeting Induced Local Lesions in Genomes) eljárás kis genetikai
varianciát mutató tulajdonságoknál kiváló módszer, amely a mutáns
populációban pont a mutációk azonosítására alkalmas. Az olasz Tuscia
Egyetem kutatóival együtt végzett vizsgálatokban sikerült waxy és
magas amilóztartalmú mutánsokat azonosítani (Sestili et al., 2010).
Nemesítési előnye, hogy az eredeti fajta introgresszió révén egy
tulajdonságra hatékonyan javítható a mutáns gén bevitelével.
Az új molekuláris nemesítési eljárások
felhasználása mellett nem lehet említés nélkül hagyni a
géntechnológiai eljárások növénynemesítési alkalmazását, a
génmódosított növényfajták előállítását.
Egyértelműen megállapítható, hogy világszerte nagy
kapacitással történik agronómiailag hasznos gének izolálása és
felhasználása transzgénikus és ciszgénikus növények nemesítésére. A
martonvásári nemesítők a genetikailag módosított búza vizsgálatát
először együttműködésben végezték a rothamstedi kutatókkal (Rakszegi
et al., 2005), majd a kutatások kiszélesedtek (2.
táblázat). A nemesítési felhasználást célzó kísérletek
2010-ben befejeződtek. Fennáll a veszélye annak, hogy lemaradásunk
olyan mértékű lesz egy sikeres külföldi áttörő eredmény gyakorlati
bevezetésekor, hogy a jövőben behozhatatlan versenyhátrányba
kerülhetünk.
Kulcsszavak: búza, előnemesítés, génbank, markertechnológia,
bioaktív komponensek
IRODALOM
Allard, Robert W. (1996): Genetic Basis of
the Evolution of Adaptedness in Plants. Euphytica. 92, 1–11. DOI:
10.1007/BF00022822 •
WEBCÍM
Arundel, Anthony – Hocke, M. – Tait, J.
(2000): How Important Is Genetic Engineering to European Seed Firms?
Nature Biotechnology. 18, 578. DOI: 10.1038/76319
Braun, Hans J. – Ekiz, H. – Eser, V. –
Keser, M. – Ketata, H. – Marcucci, G. – Morgounov, A. – Nercirci, N.
(1998): Breeding Priorities of Winter Wheat Programs. In: Braun,
Hans J. et al. (eds.) Wheat: Prospects for Global Improvement.
Kluwers Academic Publishers, 553–560.
DePauw, Ron M. – Clarke, J. M. – McCaig,
T. N. – Townley-Smith, T. F. (1998): Opportunities for the
Improvement of Western Canadian Wheat Protein Concentration, Grain
Yield and Quality through Plant Breeding. In: Fowler, D.
Brian – Geddes, W. E. – Johnston, A. M. –
Preston, K. R. (eds.): Wheat Protein Production and Marketing.
Proceedings if the Wheat Protein Symposium. Saskatoon, Saskatchewan,
Canada, 75–93.
Rabinovich, Svetlana V. (1998): Importance
of Wheat-Rye Translocations for Breeding Modern Cultivars of
Triticum aestivum L. Euphytica. 100, 323–340. DOI:
10.1023/A:1018361819215 •
WEBCÍM
Rakszegi Mariann – Békés F. – Láng L. –
Tamás L. – Shewry, P. R. – Bedő Z. (2005): Technological Quality of
Transgenic Wheat Expressing an Increased Amount of a HMW Glutenin
Subunit. J. of Cereal Science. 42, 1, 15–23. DOI:
10.1016/j.jcs.2005.02.006
Rakszegi Mariann – Boglárka N. – Kisgyörgy
N. B. – Kiss T. – Sestili, F. – Láng L. – Lafiandra, D. – Bedő Z.
(2014): Breeding of High-amylose Wheat Genotypes. Starch/Stärke. (in
press)
Sestili, Francesco – Botticella, E. – Bedő
Z. – Phillips, A. – Lafiandra, D. (2010): Production of Novel
Allelic Variation for Genes Involved in Starch Biosynthesis through
Mutagenesis. Molecular Breeding. 25, 145–154. DOI:
10.1007/s11032-009-9314-7 •
WEBCÍM
Veisz Ottó – Harnos N. – Szunics L. –
Tischner T. (1996): Overwintering of Winter Cereals in Hungary in
the Case of Global Warming. Euphytica. 92, 249–253. DOI:
10.1007/BF00022852 •
WEBCÍM
Vida Gyula – Gál M. – Uhrin A. – Veisz O.
– Syed, N. H. – Flavell, A. J. – Wang, Z. – Bedő Z. (2009):
Molecular Markers for the Identification of Resistance Genes and
Marker-assisted Selection in Breeding Wheat for Leaf Rust
Resistance. Euphytica. 170, 67–76. DOI: 10.1007/s10681-009-9945-0 •
WEBCÍM
|