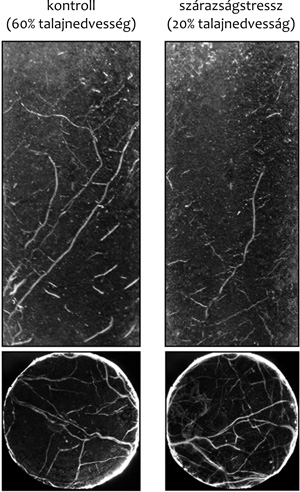
|
végeztek kísérleteket (Rajki, 1985). Beszámoltak a
Lutescens 62 tavaszi búza őszivé válásáról, miután három évben
ősszel vetették el az egyes növények utódvonalait. Ezeket a
kísérleteket erősen kritizálták. Ennek ellenére Rajki Sándor
kitartott a szerzett tulajdonságok öröklődését igazoló kísérleteinek
eredményei mellett. Az akkori kutatásokhoz kapcsolódó viták
szempontjai nyilván ma már nem tisztázhatók, de az tény, hogy a
legújabb molekuláris eredmények és módszerek, valamint az
epigenetikai ismereteink birtokában szükség lenne precíz
kísérletekkel újra elemezni az akkori állításokat, még úgy is, ha
világosan láthatóak a korábbi megközelítések korlátai. A búzában
található jarovizációs és hidegtűrési gének közül a VRN1-gén
vernalizáció nélkül alacsony kifejeződést mutat, de hidegkezelés
hatására aktiválódik, kikapcsolja a virágzást gátló VRN2-gént, és
ezzel lehetővé válik a virágzást elindító FT (VRN3) működése. A
vernalizációhoz kapcsolható epigenetikus memória a gabonafélékben a
hiszton H3-fehérjében a lizin aminosav metilációjával, és ezen
keresztül a VRN1-gén aktív kromatinszerkezetének biztosításával
szabályozott (összefoglaló: Achrem et al., 2012). A hidegkezeléssel
végzett vernalizáció továbbá a DNS metilcsoportjainak elvesztését
okozhatja, ami szintén a virágzás indukciójához vezethet (Sherman –
Talbert, 2002). Mindezek alapján igazoltnak kell tekintenünk, hogy a
vernalizáció epigenetikus variációt hoz létre, aminek öröklődése
feltételezhető, de nem történt meg a molekuláris adatok
összekapcsolása az őszi-tavaszi átmenettel (Li – Liu, 2010).
Elgondolkodtató, hogy a Gregor Mendel–Thomas Hunt Morgan és a
Micsurin–Liszenko által képviselt genetikai irányzatok vitájában
miért éppen a jarovizációhoz köthető ősziesítést használták a
szerzett tulajdonságok öröklődésének igazolására, azt a
tulajdonságot, amelyről sok évtized után beigazolódott, hogy
epigenetikus szabályozás alatt áll.
4. Az energianövények nemesítése
a kromoszómakészletük megsokszorozásával
Földünk energiaháztartásának központi szereplői a fotoszintetizáló
növények, hiszen a Nap energiájának hasznosítása révén ezek a
szervezetek a légköri CO2-gázt kémiai energiává
alakítják, és ebből állítanak elő elsődleges, megújuló szerves
anyagokat. Már napjainkban is tanúi lehetünk a megújuló biomassza
alapú energiaforrások felértékelődésének, ami számos biológiai és
műszaki probléma megoldását teszi szükségessé. Az Európai Unió
huszonhét országában a biomasszából nyert energia évenkénti
felhasználása 2020-ra több mint két és félszeresére (10 EJ) nő a
2005. évi értékhez (3,8 EJ) képest. A bioenergia-ipar számára
nyersanyagként az erdészeti fakitermelés, az energianövények
termesztése és a mezőgazdasági és kommunális hulladékok szolgálnak.
Az energetikai faültetvények sorában a rövid vágásfordulójú
energianövények jelentőségét mutatja, hogy a jelenlegi 1%-os
részesedésüket az energianövények között 2020-ra 17%-ra kívánják
növelni az European Environment Agency (Európai Környezetvédelmi
Ügynökség) 2013. évi adatai szerint. Magyarországon ennek a művelési
módnak a fehér akác (Robinia pseudoacacia L.), a nyár (Populus sp.),
valamint a fűz (Salix sp.) felel meg elsősorban (Gyurica, 2010).
Mint minden gazdasági növényünk esetében, a fafajok biológiai
teljesítőképességének maximálásában is szerepet játszik a
termőhelynek megfelelő faj megválasztása, a növénynevelés
technológiájának optimalizálása, illetve a használt fajták
tulajdonságainak nemesítéssel történő folyamatos javítása.
Tekintettel az erdészeti genetika nyújtotta elméleti alapokra,
amikor a fenotípusos bélyegek, elsősorban a biomassza hozama alapján
klónszelekciót végeznek, akkor az energiafák nemesítésében a
természetes populációk genetikai variabilitását hasznosítják (Karp
et al., 2011). A fajon belüli és a fajok közötti hibridizáció széles
körűen használt eljárás a fafajták előállításban. A kromoszómákon
térképezett DNS-szekvenciamarkerek segítségével meghatározhatók azok
a kromoszómarégiók (QTLs), amelyek kitüntetett szerepet játszanak
egyes növényi tulajdonságok meghatározásban, és amelyek segítségével
növelhető a szelekció eredményessége. A Salicaceae családhoz tartozó
két energianövény, a nyár (Populus) és a fűz (Salix) részletes
felbontású genetikai térképének összehasonlítása nagyfokú egyezést
tárt fel. A nyár (Populus trichocarpa) genomjának szekvenálását
követően, 2006 óta, a genetikai és genomikai ismeretanyag új
korszakot nyitott a fás növények biológiájában, és ezáltal az
erdészeti tudományokban (Wullschleger et al., 2014). Ez a tudományos
eredmény sokban segítette olyan gének azonosítását, amelyek fontos
morfológiai és élettani tulajdonságok kialakulását irányítják. Az új
ismeretek birtokában nincs abban kivetnivaló, hogy az előnyt
biztosító géneket a géntechnológia eszközeivel visszaépítik a nyár
genomjába. Példaként említhető az a génnemesített nyár, amelyben a
lignintartalmat csökkentették, és ezzel megnőtt az ilyen fából
nyerhető alkohol mennyisége (Van Acker et al., 2014). Bármennyire
fontosak az ilyen géntechnológiai fejlesztések, a GMO-ellenesség
közepette előtérbe kerülnek az alternatív technológiák. Így tanúi
lehetünk a kromoszómakészlet megsokszorozásával, az indukált
autopoliploidizációval történő genomformálás reneszánszának. Ez a
genetikai módszer különösen a vegetatív úton szaporított
energianövények nemesítésében nyerhet jelentőséget.
A fajra jellemző diploid (2n) kromoszómaszám
stabilan öröklődik, csak sejtosztódási rendellenességek folytán
sokszorozódhat meg a kromoszóma-állomány, és jöhet létre
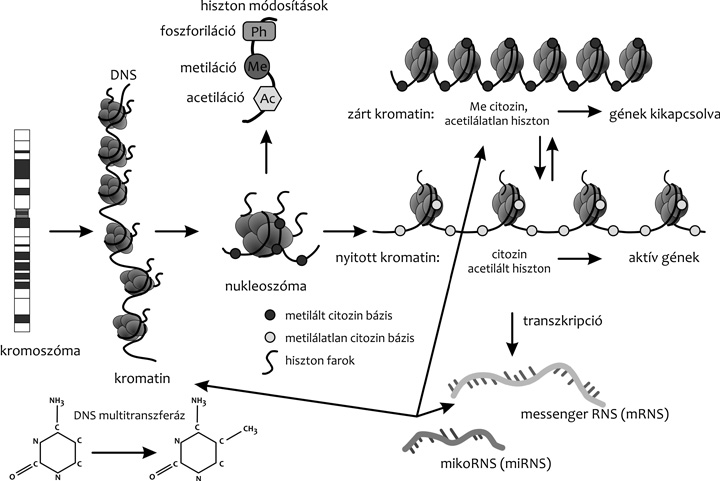
autopoliploid növény. A DNS-molekulák szintézisét követően
szerveződnek meg a nukleoszómák (2. ábra), majd az azokból
kialakuló kromatin kondenzációjával a kromoszómák. Az anyasejt
osztódásakor, a mitózis anafázisában kialakuló mikrotubulusok
biztosítják a megduplázódott kromoszóma-garnitúra szétválását a
leánysejtekben. Ha gátló vegyületek, például kolchicin hatására
sérülést szenved a mikrotubulosok funkciója, akkor nem történik meg
a kromoszómák szétválása, és megkétszereződött (4n) kromoszómaszámú
sejtek alakulhatnak ki. Ez a folyamat lejátszódhat a merisztémák
testi sejtjeiben is, ahol a 4n méretű genommal rendelkező sejtekből
új, tetraploid hajtások, növények fejlődhetnek ki. A zavar
megtörténhet az ivarsejtek képződése során azzal, hogy elmarad a
számcsökkentő, meiotikus osztódás, és a 2n kromoszómakészletet
hordozó ivarsejtek vesznek részt a megtermékenyítésben, amiből
szintén tetraploid (4n) egyedek származhatnak.
A természetben előforduló poliploid növények
meglepő tulajdonságaira Herman Nilsson-Ehle hívta fel a figyelmet
1936-ban, amikor egy óriás triploid rezgőnyárt (Populus tremula)
talált. A poliploidia gyakori a természetes növénytársulásokban, és
szerepet játszik az alkalmazkodásban. A zárvatermő növények
50–70%-a, többek között számos termesztett faj is megsokszorozódott
genommal rendelkezik. A mesterséges autopoliploidizáció mint
nemesítési módszer régi és sikeres múlttal rendelkezik. Albert
Francis Blakeslee és Amos Geer Avery 1937-ben megjelent
közleményével gyakorlatilag egy időben számolt be a tihanyi
Biológiai Kutatóintézetből Győrffy Barna a kolchicin kezeléssel
előállított poliplod növényekről, például a tetraploid paprikáról
(1938). Győrffy Barna aktív szerepet vállalt a poliploid cukorrépák
előállításában Sopronhorpácson, ahol Sedlmayr Kurt vezetésével igen
sikeres nemesítési programot valósítottak meg. Ennek során a legjobb
fajtáknak a diploid × tetraploid keresztezéssel előállított
triploidok bizonyultak. Ez a megközelítés eredményes az alma, a
körte vagy akár a dinnye nemesítésében.
Az erdészeti fajok nemesítésében is előnnyel
hasznosíthatók a megnövekedett kromoszómaszámmal rendelkező
genotípusok, amelyek megtalálhatók a természeti populációkban,
illetve mesterségesen is előállíthatók. A rövid vágásfordulóval
termesztett energiacserjék nemesítésében is előtérbe került a
poliploidizáció, tekintettel a kedvező tulajdonságok kialakításának
lehetőségére. Tetraploid nyár növényeket sikeresen állítottak elő in
vitro tenyészetekben úgy, hogy levelekből regeneráltak hajtásokat,
és azokat kolchicin-oldattal kezelték (Cai – Kang, 2011). Másik
megközelítésként a csíranövények kolchicin-kezelése is sikeres volt
a tetraploid akác előállítására (Harbard et al., 2012). Az
autotetraploid energiafűz változatok, amelyeket az in vitro
szaporított növények hónaljrügyeinek kolchicinezésével sikerült
előállítani, szintén több figyelemre méltó tulajdonsággal
rendelkeznek a diploid növényekkel való összehasonlításban. A
4. ábrán látható, hogy a
megduplázott DNS-állománnyal, illetve 2n=76 kromoszómával rendelkező
tetraploid energiafüzek levelei szélesebbek és nagyobbak, mint a
diploid változaté (Dudits et al., nem közölt).
A mesterséges autotetraploid energianövények
jellemzése nagyobb méretű légzőnyílásokat, megnövekedett CO2-megkötést,
illetve hatékonyabb fotoszintézist mutatott ki. Külön jelentősége
van annak, hogy ezek a növények jobban tudnak alkalmazkodni a
szélsőséges környezethez. Ismert a tetraploid akácnövények sótűrése.
Ami a poliploid fafajok növekedési intenzitását illeti, fajonként,
genotípusonként különböző adatokat találunk. Így az előállított
só-stressztoleráns akác esetében nem találtak különbséget a diploid
és tetraploid növények hajtásnövekedésében. Ezzel szemben a
PolygenomeX ausztrál biotechnológiai cég olyan tetraploid császárfát
(Paulownia tomentosa) állított elő, amely lényegesen gyorsabban
növekedett, és nagyobb fahozamot mutatott. Külön érdekességük
ezeknek a növényeknek, hogy kromoszómakészletüket titokban tartott
epigenetikai módszerrel sokszorozták meg (URL1). A nyírfa (Betula
platyphylla) tetraploid–diploid összehasonlításakor az
RNS-szekvenálással végzett génkifejeződési vizsgálat több ezer gén
esetében tárt fel eltérést. Az auxin- és etilénszintézist, illetve a
jelátviteli utakat befolyásoló géneknél lehetett a fenotípusos
változásokat összekötni a génkifejeződési paraméterekkel (Mu et al.
2013). Az autoploiploid energianövények eddig megismert kedvező
tulajdonságai elég alapot nyújtanak a kutatás kiszélesítéséhez, és
megalapozzák az üzemi értékeléseket.
5. Feltárulnak a növények életének titkai,
hogy elég élelmiszer, energia
és élhető környezet lehessen
A növényekkel kísérletező kutatót igen gyakran a megismerés
szenvedélye hajtja, amikor meg akar érteni egy-egy biológiai
folyamatot, a növények viselkedését. Valóban számos izgalmas esemény
történik a csírázástól a vegetatív szervek növekedésén át az
ivarszervek kialakulásáig, a megtermékenyítésig, amivel egy új
életciklus veszi kezdetét. A növénytudomány sem kivétel abban, hogy
a felfedezés élményén túl ismereteinket sokban bővítik azok az
eredmények, amelyek célorientált ipari fejlesztések során születnek.
Összességében lenyűgöző az a hatalmas előrehaladás, amelynek a
legutóbbi évtizedekben tanúi lehettünk. Ebben nagy része van a
géntechnológiának, majd a genomikának és a különböző „-omikáknak”.
Természetesen a technológiai háttér is lényegesen korszerűsödött.
Mind több növény esetében tárult fel a DNS-szekvenciában kódolt
genetikai információ, mutánsgyűjtemények és transzgenikus (GM)
növények segítik a génfunkciók megismerését. Teljesült az elvárás, a
növényeknél is lehetséges a génspecifikus mutáció vagy génbeépítés
megvalósítása. A részletes kromoszómatérképek alapján azonosíthatók
a genetikai szabályozás funkcionális egységei, javítható a
szelekciós munka eredményessége a nemesítésben. A genomika rohamos
fejlődése szükségessé tette a fenomikai fejlesztéseket (Dudits,
2012). Hatalmas kísérleti adatbázisok használhatók a növényi
funkciók modellezéséhez. Mint komplex rendszereket kezelhetjük a
fejlődési programot, a szervek növekedésének, differenciálódásának
szabályozását vagy akár a környezeti tényezőkhöz történő
alkalmazkodást. Feltárulnak azok a szignálátviteli utak, amelyek a
kórokozókkal, kártevőkkel szembeni védelmet biztosítják. Az
ökológiai szemléletet is átformálják a molekuláris biológia
eredményei.
Nem túlzás a növénytudományok virágkoráról
beszélni, ami természetesen a nemesítés sikereiben, az egészséget
védő és javító élelmiszerek kifejlesztésében, a megújuló zöldenergia
térnyerésében is megjelenik. Az optimista képet beárnyékolja az a
tény, hogy a növénytudományokat milyen súlyosan korlátozza a
géntechnológia-ellenesség. Győzelmét üli a tudománytalan propaganda,
ami hathatós politikai támogatással bír mind a törvényalkotásban,
mind a médiában. Az embereket folyamatosan a tényeket meghazudtoló
nézetekkel bombázzák, és így sikerül az egész társadalmat
megtéveszteni. Európa vesztésre áll ebben a technológiai versenyben,
de különösen súlyos a helyzet Magyarországon, ahol a géntechnológiai
tevékenység könnyen minősíthető akár börtönnel járó
bűncselekményként. Így nem meglepő, hogy csökken a fiatal kutatói
generáció bizalma, érdeklődése a növénytudományok iránt. Mindez
sokban veszélyezteti a magyar növénytudományok korábbi nemzetközi
hírnevének jövőjét. Csak abban bízhatunk, hogy a világtendenciák és
jól felfogott gazdasági érdekeink ki fogják kényszeríteni a mostani
elhibázott politika feladását, hogy a magyar kutatói közösség is
hatékonyan hozzájárulhasson a biológiai és a mezőgazdasági
tudományok fontos kutatási és fejlesztési eredményeihez.
Kulcsszavak: fotoszintézis, gyökérfejlődés, heterózis,
kromatinstruktúra, zöld energia, rövid vágásfordulójú cserjék,
autopoliploidizáció, géntechnológia, génnemesített növények
IRODALOM
Achrem, Magdalena – Skuza, L. – Kalinka,
A. et al. (2012): Role of Epigenetic Mechanisms in Plant Response to
Low Temperature. Acta Biologica Cracoviensia Series Botanica. 54, 1,
7–15. DOI: 10.2478/v10182-012-0014-y •
WEBCÍM
Bálint Andor (1964): Az öröklődés és
származástan alapjai. Mezőgazdasági, Budapest
Cai, Xiao – Kang, Xiang-Yang (2011): In
vitro Tetraploid Induction from Leaf Explants of Populus
pseudo-simonii Kitag. Plant Cell Reports. 30, 1771–1778. DOI:
10.1007/s00299-011-1085-z •
WEBCÍM
Chen, Z. Jeffrey (2013): Genomic and
Epigenetic Insights into the Molecular Bases of Heterosis. Nature
Reviews Genetics. 14, 471–482. DOI:10.1038/nrg3503
Dudits Dénes (2012): Genomikát és
fenomikát integráló növénynemesítés a termésbiztonságért. Magyar
Tudomány. 8, 913–922. •
WEBCÍM
Gómez-Díaz, Elena – Jordá, M. – Peinado,
M. A. et al. (2012): Epigenetics of Host–Pathogen Interactions: The
Road Ahead and the Road Behind. PLOS Pathogens. 8, 11, e1003007.
DOI:10.1371/journal.ppat.1003007 •
WEBCÍM
Gowik, Udo – Westhoff, Peter (2011): The
Path from C3 to C4 Photosynthesis1. Plant Physiology. 155, 56–63.
DOI: 10.1104/pp.110.165308 •
WEBCÍM
Guo, Mei – Rupe, M. A. – Dieter, J. A. et
al. (2010): Cell Number Regulator1 Affects Plant and Organ Size in
Maize: Implications for Crop Yield Enhancement and Heterosis. Plant
Cell. 22, 1057–1073. DOI: 10.1105/tpc.109.073676 •
WEBCÍM
Győrffy Barna (1938): Durch
Kolchizinbehandlung erzeugte polyploide Pflanzen. Die
Naturwissenschaften, 26, 33, 547. DOI: 10.1007/BF01675501
Gyurica Csaba (2010): Energetikai
faültetvény létesítésére alkalmas fajok és fajták (1.). Agroforum.
9, 64–76. •
WEBCÍM
Harbard, Jane L. – Griffin, A. R. –
Foster, S. et al. (2012): Production of Colchicine-induced
Autotetraploids as a Basis for Sterility Breeding in Acacia mangium.
Willd. Forestry. DOI:10.1093/forestry/cps041 •
WEBCÍM
Hauser, Marie-Theres – Aufsatz, W. –
Jonak, C. et al. (2011): Transgenerational Epigenetic Inheritance in
Plants. Biochimica et Biophysica Acta. 1809, 8, 459–68. DOI:
10.1016/j.bbagrm.2011.03.007
Hill, Kristine – Porco, S. – Lobet, G. et
al. (2013): Root Systems Biology: Integrative Modeling across
Scales,from Gene Regulatory Networks to the Rhizosphere1. Plant
Physiology. 163, 1487–1503. DOI: 10.1104/pp.113.227215 •
WEBCÍM
Karp, Angela – Hanley, S. J. – Trybush, S.
O. et al. (2011): Genetic Improvement of Willow for Bioenergy and
Biofuels. Journal of Integrative Plant Biology. 53, 2, 151–165.
DOI: 10.1111/j.1744-7909. 2010.01015.x •
WEBCÍM
Kempe, Katja – Rubtsova, M. – Gils, M.
(2014): Split-gene System for Hybrid Wheat Seed Production.
Proceedings of the National Academy of Sciences of the USA. 111, 25,
9097–9102. DOI: 10.1073/pnas.1402836111
Kurek, Itzhak – Chang, T. K.– Bertain, S.
M. et al. (2007): Enhanced Thermostability of Arabidopsis Rubisco
Activase Improves Photosynthesis and Growth Rates under Moderate
Heat Stress. Plant Cell. 19, 3230–3241. DOI:
10.1105/tpc.107.054171 •
WEBCÍM
Li, Xiuju – Liu, Yongsheng (2010): The
Conversion of Spring Wheat into Winter Wheat and Vice Versa: False
Claim Or Lamarckian Inheritance. Journal of Biosciences. 35, 2,
321–325. DOI 10.1007/s12038-010-0035-1 •
WEBCÍM
Mu, Huai-Zhi – Liu, Z-J. – Lin L. et al.
(2013): Transcriptomic Analysis of Phenotypic Changes in Birch
(Betula platyphylla) Autotetraploids. International Journal of
Molecular Sciences. 14, 20299–20325. DOI: 10.3390/ijms131013012 •
WEBCÍM
Ou, Xiufang – Zhang, Y. – Xu. C. et al.
(2012): Transgenerational Inheritance of Modified DNA Methylation
Patterns and Enhanced Tolerance Induced by Heavy Metal Stress in
Rice (Oryza sativa L.). PLOS One. 7, 9, e41143. DOI:
10.1371/journal.pone.0041143 •
WEBCÍM
Rajki Sándor (1985): Autumnisation. Acta
Agronomica Academiae Scientiarum Hungaricae. 34, 3–4, 275–373.
Sherman, Jamie D. – Talbert, Luther E.
(2002): Vernalization-induced Changes of the DNA Methylation Pattern
in Winter Wheat. Genome. 45, 2, 253–260. DOI: 10.1139/g01-147
Thiemann, Alexander – Fu, J.– Seifert, F.
et al. (2014): Genome-wide Meta-analysis of Maize Heterosis Reveals
the Potential Role of Additive Gene Expression at Pericentromeric
Loci. Bio Med Central (BMC) Plant Biology. 14, 88.
DOI:10.1186/1471-2229-14-88 •
WEBCÍM
Topp, Christopher N. – Iyer-Pascuzzi, A. –
Andersond, J. T. et al. (2013): 3D Phenotyping and Quantitative
Trait Locus Mapping Identify Core Regions of the Rice Genome
Controlling Root Architecture. Proceedings of the National Academy
of Sciences of the USA. 110 , 18, E1695–E1704.
10.1073/pnas.1304354110 •
WEBCÍM
Van Acker, Rebecca – Leplé, J-C. – Aerts,
D. et al. (2014): Improved Saccharification and Ethanol Yield from
Field-grown Transgenic Poplar Deficient in Cinnamoyl-CoA Reductase.
Proceedings of the National Academy of Sciences of the USA. 111, 2,
845–850. DOI: 10.1073/pnas.1321673111 •
WEBCÍM
Wullschleger, Stan D. – Weston, D. J. –
Difazio, S. P. et al. (2014): Revisiting the Sequencing of the First
Tree Genome: Populus trichocarpa. Tree Physiology. 33, 357–364. DOI:
10.1093/treephys/tps081 •
WEBCÍM
URL1
|