|
A limnológia paradoxonja, hogy alapvető
tudásanyagát a mély és nagy tavak vizsgálatából meríti, holott a
tipikus tó kicsi és sekély. Magyarország – relatíve kis mérete
ellenére – rendelkezik „nagyvizekkel”: a folyóvizek közül a Duna és
a Tisza emelendő ki, állóvizek tekintetében pedig a Balaton, a
Fertő, a Velencei-tó és a Tisza-tó, de ennek ellenére elsöprő a kis
tavak és patakok nagy száma. E vizek tudományos kutatása sporadikus;
a „történeti adatok” használhatósága, a mintavételi, feldolgozási,
stb. stratégiák különbözősége korlátozott.
Az EU Víz Keretirányelvének (VKI) 2000-es
kibocsátása paradigmatikus fordulatot jelentett a „vízminőség”
monitorozásában. A VKI célobjektumként jelölte ki a 10 ha-nál
nagyobb tavakat és a 10 km2-nél nagyobb vízgyűjtővel rendelkező
folyóvizeket. A vízgyűjtő-gazdálkodási tervek ezek rendszeres
állapotminősítését öt biológiai minőségi elemre (makrofiton,
fitobentosz, fitoplankton, makrogerinctelenek, halak) írják elő.
Mindebből következik, hogy növekvő és akkumulálódó tudáshiánnyal
nézünk szembe mindazon élőlénycsoportok esetében, melyeket a VKI nem
kezel kiemeltként, s mindazon vízterek vonatkozásában, melyek a
fenti határértéknél kisebbek. Ráadásul mikroszkopikus fajokra a
természetvédelmi monitorozás sem terjed ki.
Hazai és nemzetközi példák sora bizonyítja, hogy a
kisvizek – sokszor ökoton jellegük miatt – különlegesen fontosak a
biodiverzitás szempontjából, s veszélyeztetettségük igen nagy.
Jellemzően (de nem kizárólag) zárt medencéjű vizek, emiatt az
időjárás, hosszú távon pedig a klíma változásaira érzékenyek, s az
egyre gyakoribbá váló extrém időjárási jelenségek dramatikus flóra-,
fauna-, közösség- és ökoszisztéma szintű változásokkal járnak. E
tanulmányban olyan kutatási eredményeket összegzünk, melyek
rámutatnak e vizek természeti értékeire, biodiverzitásuk
gazdagságára, de akár állat- és humánegészségügyi jelentőségükre is.
1. Szikes tavak
A szikes tavak zárt medencéjű tavak, amelyek olyan lefolyástalan
területen fordulnak elő, ahol a nettó evaporáció mértéke hosszú
időátlagban eléri vagy meghaladja a lehulló csapadékmennyiséget.
Egyesek szezonálisan vagy akár évekre is kiszáradhatnak, másokra
állandó vízborítottság jellemző. A sós tavak a világ veszélyeztetett
élőhelyei közé tartoznak. Számuk az elmúlt évtizedekben
megfogyatkozott, és a prognózisok szerint 2025-re tovább csökken
(Williams, 2002). A Kárpát-medencében a szikes tavak 90%-a védett
területen található, legtöbbjük a nemzeti parkjainkban, mindegyikük
a törvény erejénél fogva (ex lege) védett élőhely és egyben Natura
2000-es terület. Fizikai és kémiai tulajdonságaikat a tó
geomorfológiája, geokémiája, a párolgás és a csapadék határozza meg.
Sótartalmuk és ionösszetételük tág határok között változik. Olyan
speciális ökológiai igényű, ritka fajok élnek itt, amelyek semmilyen
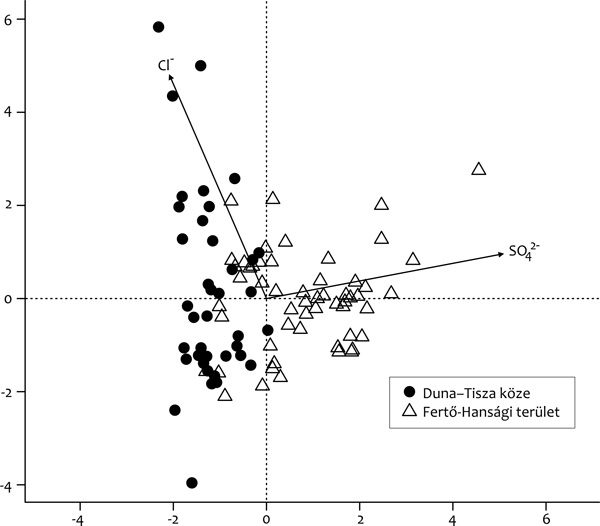
más víztípusban nem fordulnak elő. A kovaalgák nagyon jól
alkalmazható indikátorfajok, minthogy elsődlegesen olyan limnológiai
paraméterekre reagálnak, melyek a szikes tavak természetes
állapotának megőrzésében alapvető fontosságúak, ilyen például a
vezetőképesség, a HCO3-, a SO42-
és a hőmérséklet. Egészen más a kovaalga-összetétel a Duna–Tisza
közén, ahol HCO3--dominancia jellemző, mint a
Fertő–Hanság-régióban, ahol a magas HCO3-
mellett a SO42- ion mennyisége is kiemelkedő
(Stenger-Kovács et al., 2014; 1.
ábra).
A magas HCO3--tartalom jó indikátora a
Nitzschia supralitorea, míg a SO42- ioné a N.
aurariae és a N. frustulum. Hogy mely fajok kerülnek majd előnybe a
másikakkal szemben, vagy mely fajok lesznek sikeresek azáltal, hogy
a magasabb hőmérsékletet, sótartalmat kedvelik vagy tolerálják, arra
ökofiziológiai kísérletek adhatnak választ.
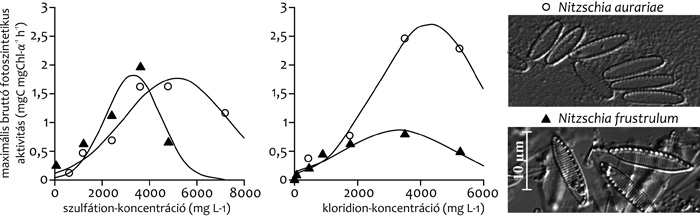
Két, törzsfejlődéstanilag és morfológiailag
hasonló, szikes tavakból kitenyésztett faj, a Nitzschia aurariae és
a N. frustulum fotoszintézisét laboratóriumi vizsgálatokban ~32,
illetve 28 °C-on találtuk optimálisnak (2.
ábra). Mindkét faj érzékeny a hidegre, fotoszintézisük 15 °C
alatt jelentősen csökken. Habár mind a két kovaalga a szikes
élőhelyek jellemző faja, megjelenésük és dominanciájuk eltérő,
amelyre magyarázatot adhat az ökológiai igényeik közötti különbség:
a fajok széles toleranciával jellemezhetők, de a N. aurariae
szulfát- (5336 mg L-1) és kloridion- (4283 mg L-1) koncentráció
optimuma nagyobb, mint a N. Frustulumé (3332 és 3253 mg L-1).
2. Szilíciumpikkelyes flagelláták
A szilíciumpikkelyes algák a plankton egy speciális csoportját
alkotják. Ökofiziológiai jellemzőjük, hogy nem rendelkeznek olyan
enzimekkel, melyek a hidrogén-karbonát szénforrásul való
felhasználását és a foszfát lúgos vizekből való felvételét lehetővé
tennék. Ostoros mozgásuk „imbolygó”, potenciálisan mixotrófok. E
jellemzők behatárolják azon élőhelyeket, melyekben előfordulnak:
enyhén savas, közepes-magas huminanyagtartalommal rendelkező,
szélhatásnak kevéssé kitett (tehát kicsi) víztestek. E víztípus
geokémiai okok folytán Magyarországon ritka, s egyes területektől
eltekintve világviszonylatban is jellemzően szigetszerű
előfordulású. Mintegy 300 faj tartozik e csoportba, köztük jelentős
számban olyanok, melyeket jól körülhatárolható cirkumpoláris,
bipoláris vagy pantropikus elterjedés jellemez. Magyarország
speciális biogeográfiai határhelyzetben van e csoport algáinak
szempontjából. A kora tavaszi időszakban a nordikus fajok jelennek
meg nagy faj- és egyedszámban az erre alkalmas kisvizekben, nyáron a
tropikus fajok előfordulása jellemző, így elterjedési mintázataik
változása klímaváltozási folyamatokat indikálhat.
A Kab-hegy bazaltkarsztmezőin számos szélvédett
tavacska található, melyek vize az alapkőzet jellege és a behulló
avar bomlása miatt huminanyagokkal színezett, enyhén savas
kémhatású. A nyári hónapokban csak a kifejezetten csapadékos években
nem száradnak ki. Természetvédelmi értéküket mutatja, hogy többet
közülük tavasszal a békaliliom (Hottonia palustris) virágszőnyege
borít. A 2013 március-áprilisában, öt tavacskából gyűjtött tizenöt
mintában elektonmikroszkópos vizsgálatokkal egyedülállóan gazdag
flórát mutattunk ki. Összesen 35 Crysophyceae fajt azonosítottunk:
egy Chyrsophaerella, egy Chrysodidymus, huszonkét Mallomonas, hat
Paraphysomonas, három Synura, egy Spiniferomonas és egy Tessellaria
fajt.
Nyolc olyan konstans fajt találtunk, amely mind az
öt tóban jelen volt (1.
táblázat; [az öt jelzetlen függőleges oszlop az 5., 4., 3.,
2. és 1. tavat jelenti], 3. ábra),
továbbá tíz olyan fajt is azonosítani tudtunk, amelyek a vázolt
élőhelytípuson belül is csupán bizonyos élőhelyekhez köthetők. Az
egyes élőhelyeken átlagosan tíz fajt azonosítottunk, közöttük
szignifikáns különbséget fajszám terén nem találtunk, viszont az
egyes élőhelyek különleges és egyedi flórával jellemezhetők.
3. Dendrotelmák
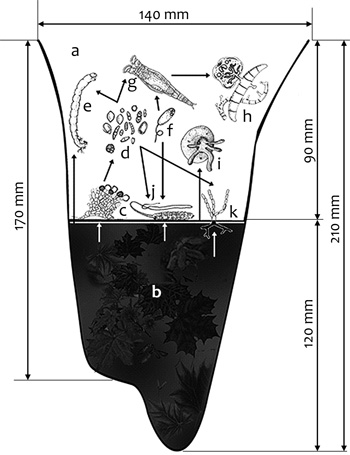
A vízi mikroökoszisztémák közé tartoznak a dendrotelmák (vízzel telt
faodvak, vízzsebek), melyek lényegében természetes úton kialakult
esővízgyűjtő élőhelyek.
Általában a fák ágelágazásainál, illetve
talajközeli gyökérrészeknél alakulnak ki, az esővizet és a
mikroorganizmusok számára jelentős táplálékforrást (avar, pollen,
termések, elpusztult rovarok) gyűjtve össze kis térfogatukban (0,1–2
liter). Ez megteremti a feltételeit annak, hogy számos
élőlénycsoport (kisméretű rákok, szúnyoglárvák, húr-, kerekes- és
fonálférgek, házatlan és házas amőbák, csillósok, ostorosok, vízi
bogárlárvák és algák) népesítse be őket
(4. ábra). A vízzel telt
faodvak humánegészségügyi szempontból is kiemelt figyelmet
érdemelnek az ott megtelepedő vektorok (például szúnyoglárvák)
miatt. A klímaváltozás eredményeképp a dendrotelmák kiszáradása és
eltűnése gyakoribb jelenséggé válhat, így kiemelt figyelmet
érdemelnek mind sérülékenységük, mind biodiverzitási értékeik miatt.
A dendrotelmák módszertani előnyeként említhető a
költséghatékony és gyors mintavétel, hiszen itt akár egy
mintavétellel is reprezentatív adatokhoz juthatunk a teljes
élőhelyről (Yanoviak – Fincke. 2005). Néhány csoport (például
makrogerinctelenek) esetén viszont ez nagy beavatkozást (károsítást)
okoz, ellehetetlenítve az időbeli vizsgálatokat. Roger L. Kitching
(2009) a módszertan széles spektrumát mutatja be, ám nélkülözve a
mikroszkopikus gombákat, melyek jelentős szerepet töltenek be a
dendrotelmák lebontási folyamataiban. Említésre méltó, hogy a vízzel
telt odvak gombaflóráját elsőképp magyar kutatók tárták fel
(Gönczöl, 1976; Gönczöl – Révay, 2003), és az ő munkáikon kívül e
témakörben alig látott szakirodalom napvilágot. Gombaközösségének
vizsgálatához nem szükséges destruktív módszert alkalmazni,
elégséges az odú egy kis vízmennyiségének vizsgálata mikroszkóposan
vagy molekuláris biológiai módszerekkel.
Egy ötéves kutatás során egy korai juharon
kialakult dendrotelmában 140 gombafajt, köztük számos ismeretlen
taxont detektáltunk, mely figyelemre méltó biológiai diverzitást
jelez. Egy jelenleg értékelés alatt álló kutatás során három további
faodú (két berkenye, egy diófa) gombaközösségét és gerinctelen
faunáját tártuk fel. Mindkét vizsgálatsorozat eredményei jelzik,
hogy az egyes faodúk más-más gombaközösséggel bírnak, valamint hogy
az időjárási paraméterek szélsőséges váltakozásai jelentős hatást
gyakorolnak a dendrotelmák közösségeire. Az
|