|
3. Pozicionális hatások és telomerek • Legújabban a
kromatinstruktúra-pozicionálás megváltozását, a sejtmagon belüli
kromatinátren-deződéseket is epigenetikai hatásoknak tulajdonítják
(Zhu et al., 2013). Egyes epigenetikai hatások a DNS-struktúra
hozzáférhetőségét, mások a rendkívül jelentős sejtélettani funkciót
ellátó telomerek hosszát befolyásolják. Ettől a szakasztól függ
ugyanis a kromoszómák stabilitása, a telomera nélküli kromoszómák
hajlamosabbak a fúzióra, ami rendellenes működéshez vezet. Másik
funkciójuk, hogy a sejtosztódáskor bekövetkező DNS-másolás során
fenntartják a lánc normális hosszát. A régió jellegzetes, guaninban
gazdag szakaszát egy telomeráz nevű enzim szintetizálja (Beery et
al., 2012). A telomerák a kromoszómák végén található, a kromoszómák
integritásának, stabilitásának megőrzésében elengedhetetlenül fontos
„kupakszerű” régiók. A telomeráknak valószínűleg szerepük lehet a
sejtmag háromdimenziós szerkezetének kialakításában is.
Legújabb eredmények szerint bizonyos környezeti
hatások (például vírusfertőzések) a genomon belül jelentős arányban
előforduló ugráló genetikai elemek (transzpozonok) aktiválódására
hatnak, tehát a genetikai elemek mobilitása is, legalább is részben,
epigenetikai kontroll alatt áll (Fedoroff, 2012).
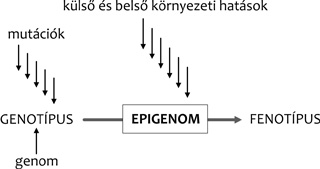
Az epigenetikai változásokat előidéző hatások
Ezek közé soroljuk az anyai hatást a magzat kihordása során, az
egyedfejlődést irányító külső faktorokat, a külső és belső környezet
tényezőit, mint amilyen a stressz, a táplálkozás, a sport, a fény, a
dohányzás, az alvásmennyiség, vagyis az életmód, valamint a
pszichoszociális környezet. Ezek mind módosíthatják a genom
aktivációját, ezért a kutatók epigenetikai tényezőknek nevezik őket.
A sokrétű anyai hatásban, az átlag negyven hetes
közvetlen anya–magzat kapcsolat során túlmenően az anya
táplálkozásán, mozgási aktivitásán és esetleges betegségeinek
hatásán túl sok más hatás érvényesül (Soubry et al., 2013). Ilyen az
anyai hang, beszéd is, amelynek jelentőségét ma már epigenetikai
mérések is igazolják. Az anyai hatás folytatódik a születés után az
érintés és illatingerek hatására is. Kiemelt epigenetikai
jelentőséget tulajdonítanak a szoptatásnak.
A táplálkozás feltételezhető epigenetikai
vonatkozásaira (Ivanova et al., 2012) utal például az egerek
szőrzetszínének öröklődésmenete, melyet a vemhes anyák táplálékának
módosításával lehet befolyásolni. Egy genisztein nevű szójaösszetevő
például a DNS-molekula egyes szakaszain hipermetilációt okoz, amely
az ivadékokban mozaikos, foltos szőrzetet eredményez, de hatására az
allergiás légúti betegségek gyakorisága is nő.
Nagyon lényeges epigenetikai tényező a mozgás, a
sport (Ehlert et al, 2013). Kiderült például, hogy a rendszeres
testmozgás gyulladásgátló hatású, mivel a vázizomzatban is
termelődik interleukin-6, amely fokozóan hat a cukor- és
zsírlebontásra.
Ami a pszichoszociális környezet epigenetikus
hatásait illeti, már végeztek vizsgálatot például olyan anyákon,
akik krónikusan beteg gyereket gondoztak. Bennük aktívabban ment
végbe a DNS-végeken elhelyezkedő telomerrégiók rövidülése, azaz a
sejtöregedés és a telomeráz a megfigyelt anyákban alacsonyabb
aktivitást mutatott a kontrollcsoporthoz képest. Ezt a jelenséget
állatmodellekben is észlelték (Liberman et al., 2012; Combs-Orme,
2013).
A jövő: személyreszabott orvoslás
A személyre szabott medicina és a tudatos egészségnevelés: a primer
prevenció (Najafzadeh et al., 2013) az epigenom szintjén történik.
A betegségek genetikai struktúrájának felderítése
óriási jelentőségű az orvoslás szempontjából, hiszen számos esetben
ugyanaz a gyógymód (vagy gyógyszer) mindenkin segít, míg gyakran
minden betegnél más beavatkozás lesz a leghatékonyabb.
A távlati célok megvalósulásáig azonban még több
kritikus problémát kell megoldani.
Először is szükség lesz még sokkal több ember
genomszekvenciájának ismeretére. 2010 végén fejeződött be az ún.
„1000 genom projekt“, ennek alapján 2012-ben publikálták ENCODE cím
alatt (Encode Project Consortium, 2012) az eredményeket. Kínai
genetikusok közeli célul tűzték ki több millió ember teljes
genomjának elolvasását. Az „új generációs szekvenálási módszerek
bekerülése és rohamosan csökkenő költségei miatt igen valószínű,
hogy ez a cél pár éven belül meg fog valósulni. Ez valóban
genomszintű asszociációs vizsgálatokat (GWAS) tesz majd lehetővé.
Másodszor, meg kell tudnunk mondani minden egyes
variánsról, hogy hozzájárul-e a betegséghez vagy például egy adott
gyógyszer lebontásának kinetikájához, és ha igen, milyen mértékben.
Ennek megállapítása igen nehéz feladatnak ígérkezik, mert a
betegségeket okozó variánsok száma valószínűleg nagy és a legtöbb
emberben különböző.
Mindazonáltal ennek a genetikai információnak a
birtokában megbecsülhető lesz majd a betegségek kialakulásának
genetikai kockázata még azok bekövetkezte előtt, ami tudatos
epigenetikai beavatkozási lehetőséget nyújthat a megelőzésre. A
primer prevenció egyik lehetősége az epigenetikai hatások
módosítása. Például az elhízásra hajlamosító genetikai variánsokat
hordozó gyerekeknél korai életmódváltoztatással a későbbi, az
elhízással fokozottabban hajlamosító megbetegedés elkerülhető lehet.
Ezen a szinten is lényeges a teljes epigenom vizsgálata (EWAS).
Végül a genetikai információra alapuló orvoslás
talán legnagyobb kihívása a variánsokat hordozó gének működését
célzottan befolyásoló és legkisebb mellékhatást előidéző
gyógyszerek, illetve személyre szabott vakcinák kifejlesztése lesz.
Nem kétséges, hogy a személyre szabott orvoslás, a
rendszerszemléletű medicina megoldandó problémái között a 21. század
orvosi genetikai/genomikai kutatásai mellett a társadalmi, szociális
és nevelési (mentális epigenetikai) kérdések is kiemelt
jelentőséggel jelentkeznek a közeljövőben.
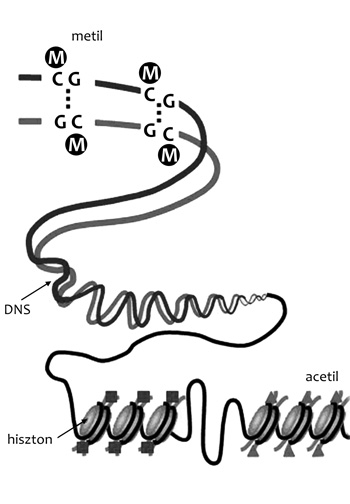
Kulcsszavak: genetika, epigenetika, DNS, metiláció, acetiláció,
mikroRNS
IRODALOM
Alberts, Bruce – Bray, D. – Lewis, J. –
Raff, M. – Roberts, K. – Watson, J. D. (eds.) (2002): The Molecular
Biology of The Cell. Fouth Edition, Garland
Beery, Annaliese K. – Lin, J. – Biddle, J.
S. – Francis, D. D. – Blackburn, E. H. – Epel, E. S. (2012): Chronic
Stress Elevates Telomerase Activity in Rats. Biology Letters. 8, 6,
1063–1066. DOI: 10.1098/rsbl. 2012.0747 •
WEBCÍM
Boros Imre M. (2012): Histone Modification
in Drosophila. Briefing in Functional Genomics. 11, 4, 319–331. DOI:
10.1093/bfgp/els029
Combs-Orme, Terri (2013): Epigenetics and
the Social Work Imperative. Soc Work. 58, 1, 23–30. DOI:
10.1093/sw/sws052
Ehlert, Tobias – Simon, P. – Moser, D. A.
(2013): Epigenetics in Sports. Sports Medicine. 43, 2, 93–110. DOI:
10.1007/s40279-012-0012-y
Encode Project Consortium (2012): An
Integrated Encyclopedia of DNA Elements in the Human Genome. Nature.
489, 7414, 57–74. DOI: 10.1038/nature11247 •
WEBCÍM
Falus András – Molnár Viktor (2008):
Closer to the Completed Unity: Messenger and MicroRNA Profiling. An
Introduction. Seminars in Cancer Biology. 18, 2, 77–78.
DOI:10.1016/j.semcancer.2008.01.006
Fedoroff, Nina V. (2012): Presidential
Address. Transposable Elements, Epigenetics, and Genome Evolution.
Science. 338, 6108, 758–767. DOI: 10.1126/science.338.6108.758 •
WEBCÍM
Hunt, Clayton R. – Ramnarain, D. –
Horikoshi, N. – Iyengar, P. – Pandita, R. K. – Shay, J. W. –
Pandita, T. K. (2013): Histone Modifications and DNA Double-strand
Break Repair after Exposure to Ionizing Radiations. Radiation
Research. 31 Jan. Epub Ahead of Print DOI: 10.1667/RR3308.2
Ivanova, Elena – Chen, J. H. –
Segonds-Pichon, A. – Ozanne, S. E. – Kelsey, G. (2012): DNA
Methylation at Differentially Methylated Regions of Imprinted
Genesis Resistant to Developmental Programming by
Maternal Nutrition. Epigenetics. 10, 1200–1210. DOI:
10.4161/epi.22141 •
WEBCÍM
Jablonka, Eva (2012): Epigenetic
Variations in Heredity and Evolution. Clinical Pharmacology &
Therapeutics. 92, 6, 683–688. DOI: 10.1038/clpt.2012.158 •
WEBCÍM
Koutsounas, Ioannis – Giaginis, C. –
Patsouris, E. – Theocharis, S. (2013): Current Evidence for Histone
Deacetylase Inhibitors in Pancreatic Cancer. World Journal of
Gastroenterology. 19, 6, 813–828. DOI: 10.3748/wjg.v19.i6.813 •
WEBCÍM
Liberman, Shayna A. – Mashoodh, R. –
Thompson, R. C. – Dolinoy, D. C. – Champagne F. A. (2012):
Concordance in Hippocampal and Fecal Nr3c1 Methylation Is Moderated
Bymaternal Behavior in the Mouse. Ecology and Evolution. 12,
3123–3131. DOI: 10.1002/ece3.416 •
WEBCÍM
Mamrut, S . – Harony, H. – Sood, R. –
Shahar-Gold, H. – Gainer, H. – Shi, Yj. – Barki-Harrington, L. –
Wagner, S. (2013): DNA Methylation of Specific CpG Sites in the
Promoter Region Regulates the Transcription of the Mouse Oxytocin
Receptor. PLOS ONE. 8, 2, E56869. DOI: 10.1371/Journal.Pone.0056869.
•
WEBCÍM
Najafzadeh, Mehdi – Davis, J. C. – Joshi,
P. – Marra, C. (2013): Barriers for Integrating Personalized
Medicine Into Clinical Practice: A Qualitative Analysis. American
Journal of Medical Genetics A. 161, 4, 758–763. DOI:
10.1002/Ajmg.A.35811
Soubry, Adelheid – Schildkraut, J. M. –
Murtha, A. – Wang, F. – Huang, Z. – Bernal, A. – Kurtzberg, J. –
Jirtle, R. L. – Murphy, S. K. – Hoyo, C. (2013): Paternal Obesity Is
Associated with IGF2 Hypomethylation in Newborns: Results From a
Newborn Epigenetics Study (NEST) Cohort. BMC Medicine. 11, 1, 29.
DOI:10.1186/1741-7015-11-29 •
WEBCÍM
Zhu, Jiang – Adli, M. – Zou, J. Y. –
Verstappen, G. – Coyne, M. – Zhang, X. – Durham, T. – Miri, M. –
Deshpande, V. – De Jager, P. L. – Bennett, D. A. – Houmard, J. A. –
Muoio, D. M. – Onder, T. T. – Camahort, R. – Cowan, C. A. A. –
Meissner, A. – Epstein, C. B. – Shoresh, N. – Bernstein, B. E.
(2013): Genome-Wide Chromatin State Transitions Associated with
Developmental and Environmental Cues. Cell. 152, 642–654. DOI:
10.1016/j.cell. 2012.12.033 •
WEBCÍM
|