Kritikus térszerkezeti elemek élőhelyhálózatokban
Jordán Ferenc
PhD, MTA–ÖBKI, Vácrátót
jordan.ferenc @ gmail.com
Bevezetés
A természetes élőhelyek beszűkülése és felaprózódása olyan problémakör, melynek megértése és kezelése a természetvédelem egyik legnagyobb kihívása. Egy faj számára a korábban összefüggő élőhelyét alkotó maradványfoltok közti átjutás a túlélés kulcsa lehet, mint ahogyan ez már egyre több faj esetében sejthető is. A kis foltokba szoruló lokális populációk ugyanis ennek hiányában könnyen kipusztulhatnak, genetikai és demográfiai okok miatt, valamint véletlenszerű katasztrófák hatására. A foltok között megmaradó területek olykor azonban lehetővé teszik az egyedek lokális populációk közti átjárását, ilyenkor funkcionális értelemben ökológiai folyosóként szolgálnak, még ha topográfiai értelemben nem lineáris jellegűek is. Az átjutó egyedek egyben átjutó alléleket is jelenthetnek, a genetikai változatosság fenntartása pedig a kis populációk veszélyeztetettségét csökkenti. Ha jól körülhatárolható élőhelyfoltok és jól azonosítható, funkcionáló ökológiai folyosók jellemzik egy faj élőhelyének szerkezetét, akkor érdemes a faj élőhelyhálózatáról (habitat network) beszélnünk, illetve annak tulajdonságait kutatva alaposabban értékelhetjük is a vizsgált faj túlélésének esélyeit. Meg kell azonban jegyezni, hogy a foltos élőhely önmagában nem utal mindig fragmentációs folyamatra, sok élőlény eleve ilyen térbeli előfordulást mutat.
Élőhelyhálózatok elemzése
A hálózatok manapság robbanásszerűen fejlődő tudománya sok olyan fogalmat és módszert szolgáltat az ökológusok számára is, melyek használata, illetve alkalmazása csábító, de sokszor nehezebb a vártnál. A hálózatelemzés ugyan rendkívül sokoldalú és szellemes megközelítést kínál például a génáramlási mintázatok jellemzésére, de sokszor nem több elcsépelt divatszónál, mely a kutatási prioritásoknak megfelelő védernyőt borít a legkülönfélébb kutatások fölé. Sok esetben valójában nincs is szó semmilyen hálózatról, inkább csak sok, egyszerre vizsgált objektumról, s a rendszer bonyolultsága látszólag meg is indokolja a hálózatos interpretálást. Ilyen értelemben maga az „ökológiai hálózat” kifejezés is, ha nincs pontosabban definiálva, értelmetlen és sokszor teljesen félrevezető.
A funkcionális értelemben vett ökológiai hálózatok, melyek gráfélei valóban releváns relációkat jelentenek a gráfpontok által reprezentált élőhelyfoltok között, mint például az i és a j folt közötti átjárhatóság, viszont valóban fontos kutatási objektumok. A tájökológiai kontextusban értelmezhető élőhelyszerkezet topológiai és topográfiai vizsgálata, az egyes térszerkezeti elemek (foltok és folyosók – most teljesen figyelmen kívül hagyjuk az „ugródeszkák” fogalmát) kvantitatív jellemzése, vagy a metapopulációs modellek térben explicit vizsgálata izgalmas kutatási területe az ökológiának és fontos problémaköre a természetvédelmi biológiának.
Természetes, hogy a különböző méretű, fiziológiájú, ökológiájú fajok különbözőképpen „látják” környezetüket, különböző tér- és időskálán zajló folyamatok részesei. Egy ugróvillás és a hóbagoly, egy tengeri ágascsápú rák és egy föld alatt élő fonálféreg, vagy éppen egy kérész és egy teknős számára mást jelent a táj, más élőhelyfoltok és folyosók jellemzik élőhelyüket, különböző módon érzékelik az élő és élettelen környezet változásait. Ennek megfelelően kicsit túlegyszerűsített az a nézet, miszerint a tájökológia a néhány tucat, illetve néhány száz négyzetkilométeres („a tájban zajló”) ökológiai folyamatok ökológiája. Persze itt az emberi léptékben értelmezett táj skálája erősen kitüntetett. A tájökológia skálafüggetlen tudományterület, mely sokkal inkább szemléletet ad, mintsem tematizál. Lényege talán a különböző ökoszisztémákban zajló folyamatok összekapcsolódását, illetve az ily módon jelentkező újabb típusú problémákat (például szubszidizáció, szegélyhatás, fragmentáció) nem kellemetlen zajnak, hanem megértendő és kutatható problémaként tekintő szemléletmód. Mindez pedig megérintette már a tengerbiológiát is, s manapság egyre többet tudunk a világtengerek emberi tevékenység okozta, túlhalászatra visszavezethető foltosodásáról (vö. top-predator hotspots).
Tekintsük végig az élőhelyhálózatok vizsgálatának egy hierarchikus sorát, mely egyre komplexebb helyzeteket vonultat fel, de lényegében azonos alapprobléma kapcsán: hogyan lehet kvantitatív módon meghatározni a térszerkezeti elemek relatív jelentőségét, illetve mindez hogyan segítheti a gyakorlati természetvédelmet.
Egy-egy fajra vonatkozó modellek
A különböző fajok élőhelyszerkezetének vizsgálata lényegesen eltérő nehézségű problémákat vet fel. Sok esetben sem az élőhelyfoltok, sem a köztük történő átjárást biztosító ökológiai folyosók meghatározása nem egyszerű. A mátrix területek (sem folt, sem folyosó) használhatósága, illetve a mátrix – folyosó – folt dinamika intenzitása meghatározza, melyek a vizsgálataink szempontjából „jónak” tekinthető fajok. A viszonylag könnyen meghatározható élőhelyszerkezettel jellemezhető fajok esetében viszont lehetőségünk van kétféle megközelítésű hálózatelemzésre is. Érdemes megvizsgálni, hogy hogyan változik az élőhely génáramlás-feltételeit megteremtő összefüggősége az élőhely szerkezeti elemeinek elvesztésének hatására (Jordán et al., 2003), illetve hogy hogyan növelhető meg az összefüggőség bizonyos új térszerkezeti elemek beépítésével (ezek persze leggyakrabban folyosók [Jordán, 2000]). Az 1. ábra egy, az első eset vizsgálatára alkalmas rendszert mutat be. A hálózatelemzési technikák segítenek abban, hogy meghatározzuk a topológiai, illetve topográfiai szempontból kritikus pozíciókat, és elvben konkrét természetvédelmi ajánlások alapjául is szolgálhatnak. Az egy-egy faj élőhelyére vonatkozó ismeretekre alapozott természetvédelmi gyakorlat azonban sok esetben a különböző faj- és élőhelyvédelmi programok konfliktusához vezet, így megkerülhetetlen a fenti szempontok többfajú esetekre történő kiterjesztése.
Többfajú modellek
és metaközösségi dinamika
Több faj élőhelyszerkezetének együttes elemzése felveti azt a problémát, vajon az egyes fajok számára optimális beavatkozások között van-e a minden faj számára együttesen optimális megoldás. Ilyen típusú kérdés merül fel, amikor például emberek és velük azonos területen élő védett állatok tájhasználata eltérő, és különböző tájelemek fontosak a különböző fajok számára. Nem biztos, hogy az egyes fajok esetében jellemzően kulcsfontosságú foltok vagy folyosók egyben az optimális megoldást jelentik a fajok együttese számára (vö. például tigrisek és helyi lakosok érdekkonfliktusa indiai nemzeti parkokban: milyen elrendezésben kell erdőfoltokat kialakítani, illetve védeni annak érdekében, hogy a tigrisek is és az emberek is a lehető legjobban járjanak; Harini Nagendra, személyes közlés).
A probléma különösen akkor izgalmas, ha a vizsgált fajok közötti ökológiai kölcsönhatásokat is ismerjük, tehát kicsi, de jól definiált közösségek élőhelyszerkezetére, illetve az „abban” végbemenő metapopulációs folyamatokra vagyunk kíváncsiak. Ilyenkor az egyes fajok lokális populációjára egyrészt a foltban együtt élő többi populáció hat (például növényevők a növényekre vagy paraziták a gazdáikra), másrészt a többi foltban élő, azonos fajú lokális populációk is kifejtik hatásukat (például bevándorló egyedeken át). Egyre több jel utal arra, hogy a közösségi ökológiai és a tájökológiai folyamatok ilyen jellegű összekapcsolása nagyon fontos (Crooks – Soulé, 1999), bár kétségtelenül nem egyszerű. A különböző fajok eltérő fragmentációs érzékenysége mindenesetre sok esetben jelzi, hogy a tájökológiai folyamatok jól azonosítható mintázatok szerint torzítják a közösségek szerkezetét. Közösségeikben a fragmentációra az átlagosnál érzékenyebbnek bizonyulnak az esőerdők óriásfái (mert az erdő belsejéhez adaptálódtak, és egy élőhelyfolt szélére kerülve zavarja őket például az erős szél [Laurance et al., 2000]), az árnyéktűrő növények (mert az élőhelyfolt napos szélén vagy nem érzik jól magukat, vagy alulmaradnak a versenyben [Metzger, 2000]), a csúcsragadozók (mert nagy területigényük miatt populációik vészesen kis méretűvé válnak a felaprózódó fragmentekben [Komonen et al., 2000]), a monofág mezei lepkék (mert kisebb fragmentben rizikósabb egyetlen tápnövényre specializálódni [Steffan-Dewenter – Tscharntke, 2000]), vagy a viszonylag ritka bogarak (mert kis fragmentben nagyobb eséllyel halnak ki például véletlen jelenségek következtében [Davies et al., 2000]). Vegyük észre, hogy minden példánk éppenséggel a millennium évéből származott, érzékeltetve azt a robbanásszerű folyamatot, ahogyan a szakma hirtelen ráébredt a problémakör jelentőségére (de további példákat is találunk az 1. táblázatban, a korábbi évekből is egyet-egyet).
Mindezek szellemében egyelőre inkább stratégiai, mint taktikai modellekről érdemes beszélni, bár a bonyolultság egyszerűsíthető: nincs kizárva, hogy már néhány kiemelt fontosságú faj kiemelt jelentőségű élőhelyszerkezeti elemeit megvizsgálva is meglepően reális képet kapunk a metaközösség dinamikájáról. Ilyenkor nem feltélenül szükséges minden fajt minden élőhelyfoltján monitorozni. A funkcionális szempontok kiemelése és a terepi tapasztalat hasznosítása sok felesleges pénzt, időt takaríthat meg. A 2. ábra egy ilyen stratégiai modell kereteit mutatja be.
Összefoglalás és kitekintés
A bioszféra több sebből vérzik, de ezek közül kettő igen súlyos. Az egyik az, hogy folyamatosan vesztünk el mindörökre olyan fajokat, melyeket nem is ismerünk, kihalásuk következményeit így igazán fel sem mérhetjük. A másik, hogy egyre zsugorodnak azok az élőhelyek, melyeket ma még egyáltalán természetesnek vagy természetközelinek nevezhetünk. A két folyamat ráadásul nem független egymástól, mint ahogy arra igyekeztem a fentiekben rámutatni. A fragmentáció közösségi ökológiai hatásai, és – kicsit általánosabban – a táj- és közösségi ökológiai folyamatok integrált tanulmányozása kulcskérdés. Ha a helyzet bonyolult, le kell egyszerűsíteni, de nem úgy, hogy megkerüljük a problémát. A helyzet viszont nemcsak bonyolult, hanem komoly is: a fragmentáció hatásai esetenként csak igen nagy késéssel érvényesülnek. Ezt persze nehéz mérni, de egy friss tanulmány szerint egy fragmentációs folyamat következtében kihalásra ítélt populációknak csak a fele pusztul ki az első ötven évben (Brooks et al., 1999). Ha ez általános jelenség, akkor alátámasztja azt a sejtést, hogy a legtöbb ritka faj védelme szélmalomharc, mert ha élettani értelemben még köztünk is járnak egyedeik, ökológiai, genetikai, pláne evolúciós értelemben már kihalásról beszélhetünk. Mindez persze új megvilágításba helyezheti a természetvédelmi gyakorlat egyes elemeit. Az ökológiai folyosók és élőhelyhálózatok tulajdonságainak kutatásáról magyar nyelven, népszerűsítő stílusban is akad olvasnivaló, így van némi remény a közfigyelem felkeltésére és a kutatói utánpótlás erősítésére, amire igen nagy a szükség, látva a problémák sorát.
Köszönettel tartozom a kézirat bírálóinak hasznos megjegyzéseikért és az előadóülés szervezőjének a meghívásért, valamint Varga Zoltán professzornak hasznos tanácsaiért. Munkámat az OTKA 37726 számú pályázata és Branco Weiss Fellowship (ETH Zürich) támogatta.
Kulcsszavak: hálózat, ökológiai folyosó, élőhelyfolt, fragmentáció, metaközösségi ökológia
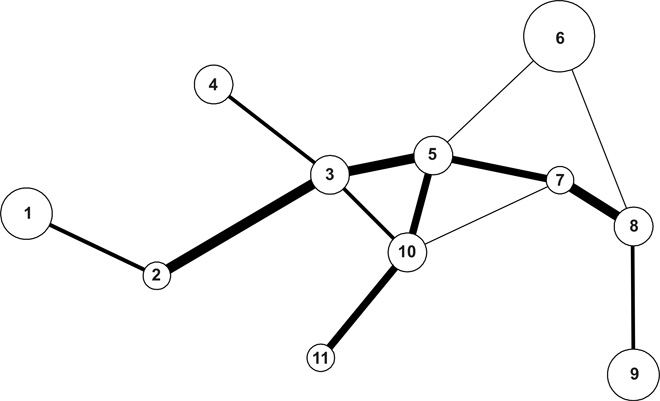
1. ábra • Az erdélyi kurtaszárnyú szöcske (Pholidoptera transsylvanica) aggteleki-karszti élőhelyhálózata. A körök élőhelyfoltokat jelölnek, sugaruk durván arányos a lokális populációmérettel. A vonalak ökológiai folyosókat reprezentálnak, vastagságuk durván arányos átjárhatóságukkal. Az egyes körökbe írt számok a foltok nevét kódolják,az alábbiak szerint: 1: Huszas-töbör, 2: Kis tisztások, 3: Szilicei kaszálók, 4: U alakú töbör, 5: Nagy Nyilas, 6: Mogyoróskúti rét, 7: Árvalányhajas, 8: Dénes-töbör, 9: Nagyoldal mögötti tisztások, 10: Gyertyánsarjas tisztás, 11: Lófej-forrás alatti tisztás (lásd Jordán et al., 2003).
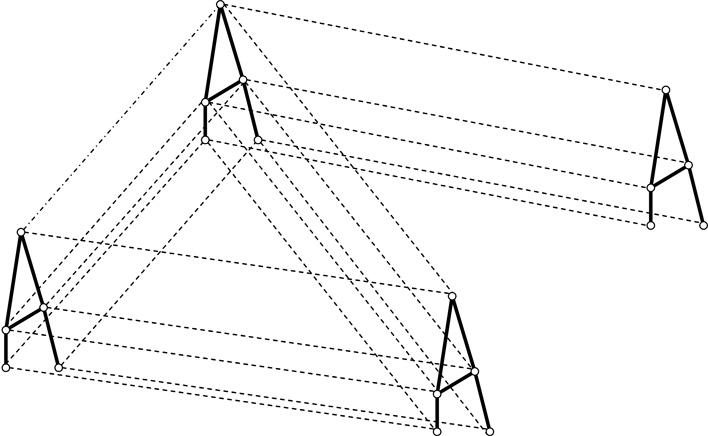
2. ábra • Egy egyszerű metaközösségi modell kereteinek illusztációja. A kis körök populációkat jelölnek, melyek között egy-egy élőhelyfoltban interspecifikus kölcsönhatások alakulnak ki (vastag vonalak), illetve a külöböző foltokon élő konspecifikus populációkat metapopulációs dinamika kötheti össze (szaggatott vonalak). Egy adott egyedet például megehet egy ragadozó (vastag vonal, közösségi ökológia) vagy átvándorolhat egy másik élőhelyfoltban élő lokális populációba (szaggatott vonal, metapopulációs, illetve tájökológia). Közösségi ökológusok és tájökológusok között sokkal intenzívebb eszmecserét kíván a különböző folyamatok természetbeli összefonódása.
Fragmentálódó Vizsgált Különösen Forrás élőhely élőlények érzékeny fajok finn erdő rovarközösség csúcsragadozó Komonen et al., 2000 német mezőgazd. lóhere fogyasztói terület és parazitoidjaik parazitoidok Kruess – Tscharntke, 1994 német gyep lepkék monofág lepkék Steffan-Dewenter – Tscharntke, 2000 német csalános rovarközösség monofág rovarok Zabel – Tscharntke, 1998 angliai mohás a moha ragadozók Gilbert et al., 1998 kövek mikroközössége ausztrál erdő 69 bogár ritka ragadozók Davies et al., 2000 kaliforniai prérik gerincesek prérifarkasok Crooks – Soulé, 1999 brazil esőerdő fák nagy fák Laurance et al., 2000 brazil esőerdő fák árnyéktűrő fajok Metzger, 2000 brazil esőerdő 993 bogárfaj ragadozók Didham et al., 1998 magyar erdőfoltok futóbogarak erdei Magura et al., 2001 élőhelyspecialisták 1. táblázat • Néhány példa a fragmentációra érzékeny fajcsoportokra. Ha a felaprózódás szelektíven érinti a különféle, együtt élő fajokat, akkor jelentősen torzulhat a közösség szerkezete, egy sor másodlagos, kaszkádszerű hatást kiváltva (a legizgalmasabb példa talán: Crooks – Soulé, 1999).
Irodalom
Brooks, Thomas M. et al. (1999): Time Lag between Deforestation and Bird Extinction in Tropical Forest Fragments. Conservation Biology. 13, 1140–1150.
Crooks, Kevin R. – Soulé, Michael E. (1999): Mesopredator Release and Avifaunal Extinctions in a Fragmented System. Nature. 400, 563–566.
Davies, Kendi F. et al. (2000): Which Traits of Species Predict Population Declines in Experimental Forest Fragments? Ecology. 81, 1450–1461.
Didham, Raphael K. et al. (1998): Trophic Structure Stability and Extinction Dynamics of Beetles (Coleoptera) in Tropical Forest Fragments. Philosophical Transactions of the Royal Society of London, Series B. 353, 437–451.
Gilbert, Francis et al. (1998): Corridors Maintain Species Richness in the Fragmented Landscapes of a Microecosystem. Proceedings of the Royal Society of London. Series B. 265, 577–582.
Jordán Ferenc (2000): A Reliability-Theory Approach to Corridor Design. Ecological Modelling. 128, 211–220.
Jordán Ferenc et al. (2003): Characterizing the Importance of Habitat Patches and Corridors in Maintaining the Landscape Connectivity of a Pholido-ptera Transsylvanica (Orthoptera) Metapopulation. Landscape Ecology. 18, 3–92.
Komonen, Atte et al. (2000): Forest Fragmentation Truncates a Food Chain Based on an Old Growth Forest Bracket Fungus. Oikos. 90, 119–126.
Kruess, Andreas – Tscharntke, Teja (1994): Habitat Fragmentation, Species Loss, and Biological Control. Science. 264, 1581–1584.
Laurance, William F. et al. (2000): Rainforest Fragmentation Kills Big Trees. Nature. 404, 836.
Magura Tibor et al. (2001): Effects of Habitat Fragmentation on Carabids in Forest Patches. Journal of Biogeography. 28, 129–138.
Metzger, Jean Paul (2000): Tree Functional Group Richness and Landscape Structure in a Brazilian Tropical Fragmented Landscape. Ecological Applications. 10, 1147–1161.
Steffan-Dewenter, Ingolf – Tscharntke, Teja (2000): Butterfly Community Structure in Fragmented Habitats. Ecology Letters. 3, 449–456.
Zabel, Jörg – Tscharntke, Teja (1998): Does Fragmentation of Urtica Habitats Affect Phytophagous and Predatory Insects Differntially? Oecologia. 116, 419–425. www.gwdg.de/~uaoe/pdf/paper/Oecologia1998,116.pdf
<-- Vissza a 2006/6 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]