|
egyik jó foltból a másikba. Számítógépes
szimulációk megmutatták, hogy ez a növekedési válasz mindenféle
környezetben kedvező, tekintet nélkül a jó foltok mennyiségére,
méretére és időbeli állandóságára (Oborny – Cain, 1997). Természetesen
ez csak azzal a feltétellel igaz, ha a foltmintázat kívülről adott,
például élettelen tárgyak vagy az adott fajnál sokkal erősebb
kompetitorok alakítják ki. Ez esetben a növény „feladata” mindössze
az, hogy hozzáigazítsa növekedésmintázatát a környezeti mintázathoz.
Más a helyzet, ha a faj aktívan befolyásolni képes a környezeti
mintázatot, például ahova belenő, onnan ki tudja szorítani a
versenytársakat. E lehetőséget is tekintetbe véve két alapstratégiát
lehet elkülöníteni: az ún. falanx és gerilla stratégiát (Lovett Doust,
L. – Lovett Doust, J., 1982).
Mint az elnevezés is mutatja, a gondolat a
hadviselésből ered; a tipikus kísérletekben azt veszik figyelembe,
hogyan reagál egy faj a kompetícióra. A gerilla – például a Trifolium
repens – elkerülő stratégiát folytat. Ha nő a kompetíciós nyomás,
növeli a térelválasztók hosszát, tehát a következő jó folt
felderítésére kezd több energiát fordítani. A falanx ezzel szemben
eleve is viszonylag rövidebb térelválasztókkal rendelkezik, és
kompetíció hatására ezeket tovább rövidíti, tehát „tömöríti” a
frontot, és fokozatosan szorítja ki az ellenfelet. Példa erre a csomós
fűfajok növekedése. A stratégiák egymáshoz képesti előnye/hátránya a
környezettől függ, például a forrás összmennyiségétől. A falanx és
gerilla tartósan is együtt élhet, ha a falanx faj olyan növénytakarót
alakít ki, melyben a gerilla számára elegendő üres folt kínálkozik
(lásd kompetíciós–kolonizációs csereviszony). A növénytársulások
szerkezetének kiépülése, stabilitása az együtt élő fajok növekedési
stratégiáinak „illeszkedésén” múlik (Oborny–Bartha, 1995).
Mennyire integrált az egyed?
Az unitér szervezetek magasan integrált lények, a moduláris
szervezetekben az integráltság foka változatos. Szélsőséges esetben
akár minden egyes modul önállóvá, a többiektől fiziológiailag
függetlenné válhat. Ezt találjuk a vegetatív szaporodású (klonális)
növényeknél, például az indával, gyöktörzzsel, gumóval szaporodó
fajoknál. Érdekes átgondolni, hogy ez esetben a genetikai értelemben
vett egyed (genet) több fiziológiai egyedből (rametből) áll. Az
individuum fogalma a moduláris élőlényeknél (még) kevésbé egyértelmű,
mint az unitéreknél (vö. Buss, 1983; Pál – Szathmáry, 2000), az adott
problémához mindig pontosan specifikálni kell, melyik fajta egyedre
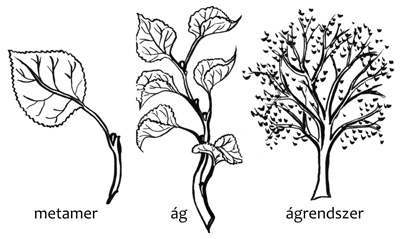
gondolunk. A legkisebb építőegység (metamer) és a teljes genetikai
egyed között (1. ábra) gyakran több, köztes hierarchiaszintet
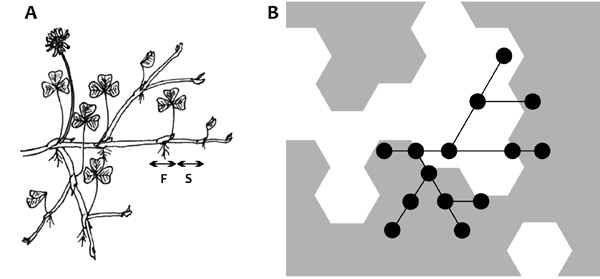
is érdemes figyelembe venni. A Trifolium repens (3.a ábra)
esete viszonylag egyszerű: itt a legkisebb építőegység, a metamer
felel meg a rametnek. Születése után egy ideig még a szülő
támogatására – víz, tápanyagok stb. transzportjára – szorul, ezután
képessé válik kifejleszteni minden olyan szervet, amely az önálló
élethez szükséges: gyökérzetet, vegetatív és reproduktív hajtást. Más
fajoknál nem a metamer, hanem egy ág vagy egy ágrendszer a ramet, a
fehér akácnál (Robinia pseudoacacia) például egy teljes fa.
Az önállósodás képessége nem feltétlenül jelent
tényleges önállósodást. Külön érdekes kérdés, hogy melyik fajnál
milyen források mozognak egyik metamerből a másikba, egyik ágból a
másikba stb., s ezeknek az áramlásoknak milyen az időbeli mintázata,
például szezonálisan vagy évek között összehasonlítva (Watson, 1986).
A kísérletezés legegyszerűbb formája a forrás radioaktív jelölése
(például a nitrogéné, foszforé). Közvetettebb, de terepen jobban
alkalmazható módszer, ha a genet különböző részeit különböző
forrásellátottságnak tesszük ki (például sok/kevés nitrogén,
fény/árnyék), és nyomon követjük a részek fejlődését. Sok fajnál az is
megtehető, hogy eközben a rametek közötti összeköttetést (például
indát) elvágjuk – a vágás miatti sérülés hatását megfelelően
kontrollálva –, s összehasonlítjuk az érintetlen esettel.
A kísérletek tanúsága szerint a növényfajok
rendkívüli változatosságot mutatnak abban a tekintetben, hogy milyen
az integráltság térbeli és időbeli mintázata (IPU – Integrated
Physiological Unit; lásd Watson, 1986). Számos fajnál azt látjuk, hogy
a rametek vagy aktívan szétválnak egymástól (például abszcíziós
szövetképzéssel), vagy összekötve maradnak ugyan, de az anyagmozgás
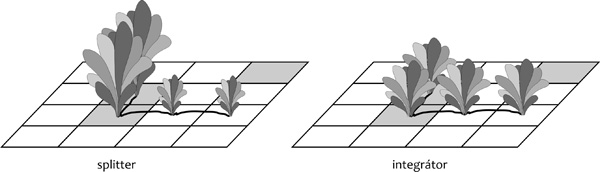
közöttük elhanyagolható. E fajok az ún. splitter stratégia képviselői.
Ellenesetben a növény részei fiziológiai összeköttetésben maradnak.
Leggyakrabban azt látjuk, hogy emiatt csökken a részek közötti
különbség, azaz az anyagmozgás az átlagolás irányába hat. Ezt gyakran
nevezik integrátor stratégiának. Megjegyzendő, hogy ritkábban az
ellenkezője is előfordul, mikor az összeköttetés hatására a részek
közötti különbségek felfokozódnak. Összességében a növény növekedése a
részek közti kompetíció és kooperáció során alakul; eközben a releváns
evolúciós egység az öröklött stratégia hordozója, a genet, tehát ennek
rátermettségét kell maximalizálni.
Munkatársaimmal – Kun Ádámmal, Meszéna Gézával, Mágori Krisztiánnal és
Ulf Dieckmannal – számítógépes szimulációk segítségével kimutattuk,
hogy az integrátor akkor kerül előnybe a splitterrel szemben, ha
viszonylag kevés a forrásban gazdag folt, ezek kicsik, és időben
változók. A splitter ugyanis a jó foltokban erősebb kompetitor, mint
az integrátor, hiszen „a gazdagnak nem kell támogatnia a szegény
rokont”. Tehát mindaddig kiszorítja az integrátort, amíg viszonylag
nagy esélye van annak, hogy jó foltból jóba lépve a
teljes jó területet vagy annak nagy részét el tudja özönleni. (A
kis, szigetszerű jó klaszterekben az integrátor már nem képes tartós
túlélésre.) Ha viszont elözönlésre nincs lehetőség, akkor az
integrátor tartósan megél a splitter által meghagyott területeken,
sőt, van a paramétertérnek olyan része is, ahol legyőzi ellenfelét. A
„szegény rokon támogatásának” ugyanis megvan az az előnye, hogy a
távolabbi jó foltokba is el lehet jutni a rossz területen átkelve (4.
ábra), sőt, a kölcsönös támogatás miatt változó foltmintázathoz is
jobban lehet alkalmazkodni. A probléma alapjában lefordítható egy
perkolációs problémára. Adaptív dinamikai modellben megmutatható, hogy
mindkét irányban lehetséges evolúciós átmenet – a splittertől az
integrátor felé és viszont –, kis mutációs lépésekkel is.
Talán a fentiekből is látszik, hogy ha a
fiziológiai integráció következményeit vizsgáljuk, ugyanúgy a
forrás-felderítés és -kiaknázás kérdése merül föl, mint a morfológiai
plaszticitás vizsgálatakor (lásd: Alpert – Stuefer, 1997, Hutchings –
Wijesinghe, 1997). A két problémakör azonban még nincs összekötve;
hiányzik egy átfogó áttekintés. Erre azért is szükség volna, mert a
két jelenség nem független egymástól: minél inkább átlagol az
integráció, annál kevésbé tud kifejeződni a morfológiai plaszticitás.
Térbeli munkamegosztás
Az [A] és [C] típusú plaszticitás egy érdekes kombinációja az ún.
térbeli munkamegosztásra ad lehetőséget. A jelenséget több fajon is
kimutatták (összefoglalásként lásd Alpert – Stuefer, 1997; Hutchings –
Wijesinghe, 1997). A tesztekben általában a növény két, egymással
összeköttetésben lévő rametjét vizsgálták. Az egyiket fényben szegény,
talajtápanyagokban gazdag helyen tartották (X ramet), a másikat éppen
fordítva, jó fényellátottságú, de gyenge tápanyag-ellátottságú helyen
(Y ramet). Ez nagyjából megfelel annak a helyzetnek, mikor a növény
egy nem túl jó vízellátottságú gyepben él, ahol egyes rametek
fűcsomókban (X) mások a fűcsomók közötti területeken nőnek (Y). A
kísérletek azt mutatták, hogy az X ramet a gyökérzetbe, az Y a hajtás
növesztésébe fektet több energiát, tehát rametszinten specializáció
történik a lokálisan bőségben lévő forrás felvételére. Ez az eredmény
éppen ellentétes azzal, amit a Liebig-törvény alapján várnánk! Az
ellentmondás az eggyel magasabb szerveződési szinten oldható fel: a
rametek megosztják egymással a forrásokat, az X → Y irányban víz és
ásványi tápanyagok, az Y → X irányban asszimilált szénhidrátok
mozognak. Az [A] és [C] típusú plaszticitás ilyen kombinációja
jelentős előnnyel járhat. Michael J. Hutching és Dushyantha K.
Wijesinghe például a kerek repkénnyel (Glechoma hederacea)
kísérletezett. Ugyanazt a talajtápanyag-mennyiséget homogén, illetve
heterogén módon osztották el a kísérleti parcellában. Utóbbi esetben
változtatták a foltméreteket. Azt találták, hogy azonos
össztápanyag-mennyiség mellett a megfelelő méretű foltokban való
elosztás akár meg is négyszerezheti a termelt biomasszát!
Konklúziók
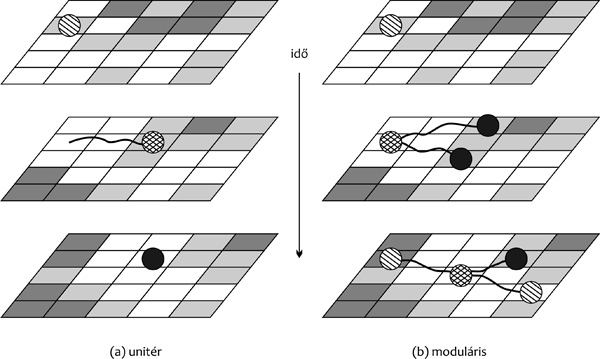
A növény számára ugyanúgy kihívást jelent a környezeti heterogenitás,
mint az (unitér) állat számára (2. ábra), az adaptív válaszok
repertoárja azonban jóval kevésbé ismert. Az bizonyos, hogy a növény –
moduláris felépítésének köszönhetően – rendelkezik néhány olyan
„trükkel”, amellyel az unitér állat nem. Először is, a növény
egyidejűleg több ponton tud mintát venni az élőhely minőségéből. (Az a
és b részábra összehasonlíthatósága végett szögezzük le, hogy
mindkettőt külön-külön úgy skálázzuk, hogy a rács celláinak mérete
akkora legyen, mint az általunk vizsgált mozgási vagy növekedési
válasz legfinomabb felbontása. Ami a növénynél néhány centiméter, az
az állatnál több méter is lehet, az alapprobléma azonban ugyanaz: a
forrásfoltok felderítése és kiaknázása.) A növény nemcsak több ponton
érzékeli a környezetet egyszerre; lokális fenotípusos plaszticitása
[A] révén arra is képes, hogy az adott (mikro)környezettől függően
más-más fenotípust alakítson ki.
A modularitás lehetőséget teremt a genetikai egyed
feldarabolására (splitter stratégia). Feldarabolódás esetén a genet
mortalitásának kockázata több, egymástól független rametmortalitás
között oszlik meg (spreading of risk; Eriksson – Jerling, 1990).
Ugyanaz a genom többféle környezetben „tesztelődik le”, ráadásul [A]
típusú plaszticitás esetén többféle fenotípus formájában.
Ha a genet nem darabolódik fel, hanem teljesen vagy
részben összekötött marad (integrátor stratégia), akkor a kockázat
szétszórása helyett megosztására van lehetőség, és kooperáció működhet
a rametek között. Ennek egyik fejlett formája az [A] és [C] típusú
plaszticitást egyesítő térbeli munkamegosztás.
Az unitér állat mozgási útvonala a növény
növekedési útvonalának felel meg (1. ábra). A [B] típusú
plaszticitás lehetőséget teremt ennek az útvonalnak az
optimalizálására. Érdekes, hogy a növény nem feltétlenül hagyja el
azokat a helyeket, amelyeken járt; ennek ütemezését a modulok
mortalitása dönti el. Múltbeli helyekről így forrás mozoghat a későbbi
helyek felé, sőt ellenirányban is.
A moduláris szerveződés következtében felfokozódik
néhány olyan evolúciós jelenség fontossága, amely az unitér
élőlényeknél is megvan. Ilyen a hierarchikus szelekció (Tuomi –
Vuorisalo, 1989), amely a növényeknél a makroszkópos szinten is
megnyilvánul (egy fa vagy egy óriásklón cserje esetén akár a több
méteres térbeli nagyságrendben is). További fontos jelenség, hogy a
szomatikus folyamatok számottevő evolúciós jelentőséget nyernek. Ez
abból ered, hogy a moduláris élőlényeknél nyílt az egyedfejlődési
program, és viszonylag késői a sejtvonalak elköteleződése, így a
(pre)szomatikus mutációk bejuthatnak a csíravonalba (Klekowski, 1988).
Különösen fontos ez a hosszú életű klónok esetén, amikor – több száz
vagy ezer év alatt – a szomatikus mutációk fellépésére számottevő
esély adódik. Ugyanemiatt jelentőssé válnak az epigenetikai
öröklődéssel átadott jellegek (lásd Jablonka – Lamb, 1995, Pál –
Szathmáry, 2000), például egy kártevő elleni „készültség” foka). A
modulpopuláció dinamikájának, a modulok össze- és szétkapcsoltságának
megértése alapvetően fontos e fajok evolúciójának megértése
szempontjából, márpedig a moduláris élőlények a Föld többsejtű
élővilágának el nem hanyagolható részét alkotják.
A tanulmány a Nest (Új Európai Iskola az Elméleti
Biológiáért és Ökológiáért) Alapítvány és az MTA támogatásával, a
szerzőnek ítélt 2009. évi Juhász-Nagy Pál-díj keretében készült.
Köszönöm továbbá az OTKA (K61534) támogatását. Hálás vagyok Fekete
Gábornak és Podani Jánosnak a kézirat átnézéséért, a javaslatokért.
Kulcsszavak: modularitás, elágazásmintázat, adaptív növekedés,
kooperáció és kompetíció, hierarchikus szelekció, darwini evolúció
IRODALOM
Alpert, Peter – Stuefer, Josef (1997):
Division of Labour in Clonal Plants. In: De Kroon, Hans – Van
Groenendael, Jan M. (eds.): The Evolution and Ecology of Clonal
Plants. Backhuys Publishers, Leiden, 137–154.
Bell, Adrian D. (1984): Dynamic
Morphology: A Contribution to Plant Population Ecology. In: Dirzo,
Rodolfo – Sarukhán, José (eds.): Perspectives on Plant Population
Ecology. Sinauer Assoc., Sunderland. 48–65.
Bell, Adrian D. (1991): Plant Form. An
Illustrated Guide to Flowering Plant Morphology. Oxford University
Press, Oxford.
Buss, Leo W. (1983): Evolution,
Development, and The Units of Selection. Proceedings of the National
Academy of Sciences of the USA – PNAS. 80, 387–1391.
WEBCÍM >
Eriksson, Ove – Jerling, Lenn (1990):
Hierarchical Selection and Risk Spreading in Clonal Plants. In: Van
Groenendael, Jan – De Kroon, Hans (eds.): Clonal Growth in Plants:
Regulation and Function. Spb Academic Publ., The Hague. 79–94.
Harper, John L. (1977): The Population
Biology of Plants. Academic Press, London.
Harper, John L. (1985): Modules, Branches,
and the Capture of Resources. In: Jackson, Jeremy B. C. – Buss, L. W.
– Cook, R. E. (eds.): Population Biology and Evolution of Clonal
Organisms. Yale University Press, New Haven. 1–33.
Hutchings, Michael J. – Wijesinghe,
Dushyantha K. (1997): Patchy Habitats, Division of Labour and Growth
Dividends in Clonal Plants. Tree. 12, 390–94.
Jablonka, Eva – Lamb, R. Marion (1995):
Epigenetic Inheritance and Evolution. Oxford University Press.
Jackson, Jeremy B. C. – Buss, L. W. –
Cook, R. E. (eds.): (1985): Population Biology and Evolution of Clonal
Organisms. Yale University Press, New Haven
Jerling, Lenn (1985): Are Plants and
Animals Alike? A Note on Evolutionary Plant Population Ecology. Oikos,
45, 150–153.
Klekowski, Edward J. (1988): Mutation,
Developmental Selection, and Plant Evolution. Columbia Univ. Press,
New York
Lovett Doust, Lesley L. – Lovett Doust,
Jon L. (1982): The Battle Strategies of Plants. New Scientist. 95,
81–84.
Molnár V. Attila (2009): Kétszáz éve
született Darwin. Az elfelejtett botanikus. Élet és Tudomány. 6,
166–169.
Novoplansky, Ariel (ed.) (2002):
Developmental Plasticity in Plants. Special Issue of Evolutionary
Ecology. Vol. 16.
Oborny Beáta – Bartha Sándor (1995):
Clonality in Plant Communities: An Overview. In: Oborny Beáta – Podani
János (eds.): Clonality in Plant Communities. Special Features in
Vegetation Science 11, Opulus Press, Uppsala. 115–127.
Oborny Beáta – Cain, Michael L. (1997):
Models of Spatial Spread and Foraging in Clonal Plants. In: De Kroon,
Hans – Van Groenendael, Jan (eds.): The Ecology and Evolution of
Clonal Plants. Backhuys Publ., Leiden, 115–127.
Pál Csaba – Szathmáry Eörs (2000): The
Concept of Fitness and Individuality Revisited. Journal of
Evolutionary Biology. 13, 348–357.
Schmid, B. (1990): Some Ecological and
Evolutionary Consequences of Modular Organization and Clonal Growth in
Plants. Evolutionary Trends in Plants. 4, 1, 25–34.
Tuomi, Juha – Vuorisalo, Timo (1989):
Hierarchical Selection in Modular Organisms. Tree. 4, 7, 209–13.
Vuorisalo, Timo – Tuomi, Juha (1986):
Unitary and Modular Organisms: Criteria For Ecological Division.
Oikos, 47, 382–385.
Watkinson, A. R. (1988): On the Growth and
Reproductive Schedules of Plants: A Modular Viewpoint. Acta Oecologica
(Oecologia Plantarum) (Ecologia Plantarum). 9, 67–81.
Watson, Maxine A. (1986): Integrated
Physiological Units in Plants. Tree, 1, 5, 119–123.
White, James (1979): The Plant As a
Metapopulation. Annual Review of Ecology and Systematics. 10, 109–145.
FÜGGELÉK
Darwin növényekkel foglalkozó könyveinek
első kiadásai:
Darwin, Charles (1862): On the Various
Contrivances by Which British and Foreign Orchids Are Fertilised by
Insects. John Murray, London
Darwin, Charles (1865): On the Movements
and Habits of Climbing Plants. The Linnean Society of London
Darwin, Charles (1875): Insectivorous
Plants. John Murray, London
Darwin, Charles (1876): The Effects of
Cross and Self Fertilisation in the Vegetable Kingdom. John Murray,
London
Darwin, Charles (1877): The
Different Forms of Flowers on Plants of the Same Species. John Murray,
London
Darwin, Charles – Darwin, Francis (1880): The Power of Movement in
Plants. John Murray, London
|