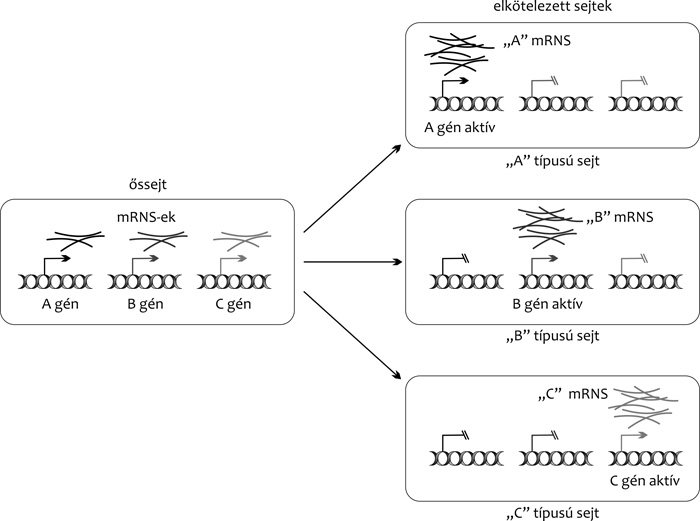
|
hozzáférhető a hírvivő RNS- (mRNS) molekulák
átírását végző apparátus számára, mint a véglegesen
differenciálódott sejtekben. Ezért számos különböző – más-más
fejlődési irány, illetve sejtfejlődési sor meghatározására képes
fehérjét kódoló – gént tudnak egyidejűleg kifejezni. (A jelenséget
„genetikai promiszkuitásnak” is nevezik) (1.
ábra) (Collas, 2009).
Potenciálisan tehát egy szöveti őssejt sokféle
genetikai program megvalósítására képes. A döntés,
hogy e lehetőségek közül adott esetben melyik realizálódik – azaz
milyen irányba kezd differenciálódni a sejt – részben valószínűségi
alapon, részben környezeti tényezők hatására történik. A valószínűség
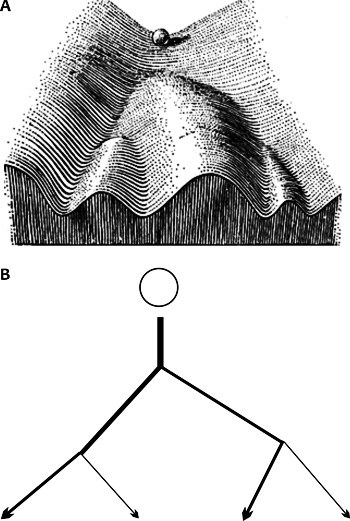
szerepét legkönnyebben a Waddington-féle „epigenetikus tájkép”
(Waddington, 1957) hasonlat segítségével érthetjük meg, ami az
őssejtben kifejeződő gének bonyolult hálózatát szemlélteti
(2. ábra). Ha egy hegycsúcsról legurítunk
egy golyót, az számos kisebb-nagyobb völgy felé gurulhat, útját
azonban igen nehéz kiszámítani. A legvalószínűbb, hogy a legmélyebb
völgy felé veszi az irányt, de ettől nagyon sok, akár egészen apró
tényező – például egy útjába kerülő kavics – is eltérítheti. Ráadásul
a völgyek elágazhatnak, szakadékban vagy újabb emelkedőben is
végződhetnek. Ugyanígy nem tudjuk biztosan megmondani, hogy egy
differenciálódásnak indult őssejtben végül is milyen
fehérjekombináció, azaz milyen megjelenési forma (fenotípus)
stabilizálódik. A szöveti őssejtek közvetlen környezetéből érkező
jelzések azonban alaposan megváltoztatják a játékszabályokat.
Igyekeznek az „őssejtgolyót” egy vagy néhány kiválasztott völgy, azaz
meghatározott sejtfejlődési sor irányába terelni. Ezeket a jelzéseket
az őssejt feldolgozza, integrálja, majd meghozza a lehetséges
döntés(eke)t – elkötelezetté válik –, és ennek megfelelően kezd
differenciálódni (Halley et al., 2008; Mohn – Schübeler, 2009).
Az érem másik oldala
A fent vázolt komplex, adaptív szabályozó rendszer – amelyben
viszonylag kis változások is több száz vagy inkább ezer gén
kifejeződését érintik (illetve érinthetik) – biztosítja a szöveti
őssejtek rendkívüli plaszticitását. Ez teszi lehetővé, hogy az
őssejtek mindkét alapvető feladatuknak megfeleljenek. Fiziológiás
körülmények között fenntartsák az adott szövet homeosztázisát, vagyis
pótolják az öregedő, pusztuló sejteket, illetve sérülés esetén
biztosítsák az érintett szövet regenerációját. Ugyanakkor a
rendszerben fellépő minimális hiba is komoly patológiás
következményekkel járhat. Az MSC-k könnyen a daganatok növekedését és
áttétek képzését elősegítő aktivált fibroblastokká vagy a különböző
létfontosságú szervek (máj, lép, vese) kötőszövetes elfajulásáért
felelős myofibroblastokká alakulhatnak.
Kulcsszavak: epigenetikus tájkép, genetikai promiszkuitás,
génexpressziós ujjlenyomat, immunszuppresszió, mesenchymalis őssejt,
myofibroblast, plaszticitás
IRODALOM
Bianco, Paolo – Robey, P. G. – Simmons,
P.J. (2008): Mesenchymal Stem Cells: Revisiting History, Concepts, and
Assays. Cell Stem Cell. 2, 313–318.
Caplan, Arnold I. (2007): Adult
Mesenchymal Stem Cells for Tissue Engineering Versus Regenerative
Medicine. Journal of Cellular Physiology. 213, 341–347.
WEBCÍM >
Collas, Phillipe (2009): Epigenetic States
in Stem Cells. Biochimica et Biophysica Acta (BBA) – General Subjects.
1790, 900–905.
Dominici, Massimo – Le Blanc, K. –
Mueller, I. et al. (2006): Minimal Criteria for Defining Multipotent
Mesenchymal Stromal Cells. The International Society for Cellular
Therapy Position Statement. Cytotherapy. 8, 315–322.
WEBCÍM >
Friedenstein, Alexander J. – Gorskaja, J.
F. – Kulagina N. N. (1976): Fibroblast Precursors in Normal and
Irradiated Mouse Hematopoietic Organs. Experimental Hematology. 4,
267-274.
Halley, Julianne D. – Winkler, D. A. –
Burden, F. R. (2008): Toward a Rosetta Stone for the Stem Cell Genome:
Stochastic Gene Expression, Network Architecture, and External
Influences. Stem Cell Research. 1, 157–168.
Jones, Ben J. – McTaggart, Steven J.
(2008): Immunosuppression by Mesenchymal Stromal Cells: From Culture
to Clinic. Experimental Hematology. 36, 733–741.
Mohn, Fabio – Schübeler, Dirk (2009): Genetics and Epigenetics:
Stability and Plasticity During Cellular Differentiation. Trends in
Genetics. 25, 129–136.
Nauta, Alma J. – Fibbe, Willem E. (2007):
Immunomodulatory Properties of Mesenchymal Stromal Cells. Blood. 110,
3499–3506.
Phinney, Donald G. – Prockop, Darwin J.
(2007): Concise Review: Mesenchymal Stem/Multipotent Stromal Cells:
The State of Transdifferentiation and Modes of Tissue Repair—Current
Views. Stem Cells. 25, 2896–902.
WEBCÍM >
Waddington, Conrad Hal (1957): The
Strategy of the Genes. Allen & Unwin, London.
|