|
pedig három – Ascl1, Brn2 és Myt1l (Vierbuchen et
al., 2010) – TF egyidejű alkalmazásával lehet alakítani. Minél
távolabb van tehát egymástól a kiindulási és az előállítani kívánt
sejttípus, annál több „mester” TF egyidejű kifejeződésére van
szükség ahhoz, hogy a transzdifferenciáció végbemenjen. Az érintett
TF-ok valószínűleg ugyanazok, amelyek az egyedfejlődés során is
biztosítják a különböző sejtfejlődési sorok elköteleződését.
A „mester” TF-okat kódoló géneket természetesen nem
csak bevinni lehet a sejtekbe, elnémításuk (génkiütéses – knock out
technika) sokszor szintén rendkívül hatékony módszer a sejtek
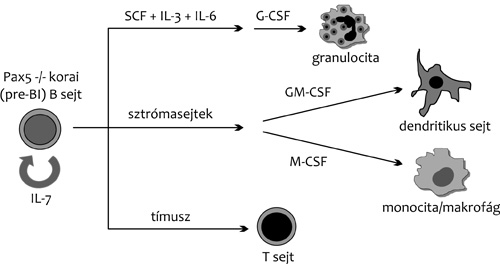
sorsának befolyásolására. A Pax5 gén például a B-limfocita fejlődési
sor mesterregulátora. Az általa kódolt Pax5 transzkripciós faktor
hiányában (Pax5 -/- knock out egerekben) a B- limfociták fejlődése a
korai (pre-BI sejt) stádiumban megáll. Az immunglobulin nehézlánc
génátrendeződés a folyamat első lépése, egy diverzitás és egy
joining szekvencia összekapcsolódása (DHJH joining) után elakad. Azt
várnánk, hogy az ilyen Pax5 -/- pre-B sejtek életképtelenek.
Ezzel szemben csontvelői sztrómasejtekre ültetve,
nagy mennyiségű IL-7 jelenlétében, önfenntartó osztódásba kezdenek
in vitro kultúrában. Ha az IL-7-et fokozatosan különböző más
citokinekkel helyettesítik a tenyészetekben, a Pax5 hiányos pre-BI
sejtekből granulociták, dendritikus sejtek és macrofágok
keletkeznek. Sőt, tímuszkultúrában vagy működő tímusz-szal
rendelkező állatba oltva T-limfocitákká is differenciálódhatnak
(4. ábra). A normális B-sejt fejlődése során tehát a Pax5
fehérje kettős szerepet játszik. Egyrészt elősegíti a
B-sejt-specifikus gének expresszióját, másrészt gátolja a többi
limfo-hematopoetikus sejtfejlődési sor „mester-regulátor” génjeinek
kifejeződését a pre-B-sejtekben (Nutt et al., 1999). Utóbbi nem a
Pax5 TF egyedi sajátsága, hanem általánosan jellemző a „mester”
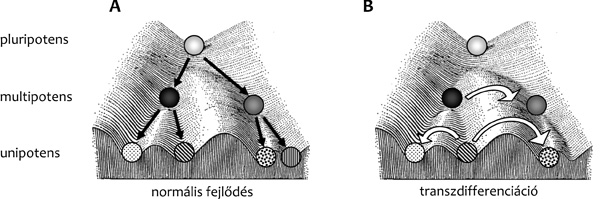
TF-okat kódoló gének működésére. A szöveti ős- és elődsejtek éppen
azért képesek még többféle genetikai program megvalósítására, mert
egyidejűleg számos „mester” TF-t fejeznek ki (azaz multipotensek,
lásd még a Bevezetőben). Amikor valamelyik „mester” TF szintje elér
egy kritikus értéket a sejtben, és az valamelyik fejlődési sor
(például: A) irányába elkötelezetté válik, akkor a
másik (B) fejlődési irány (vagy irányok) TF-ait kódoló gének
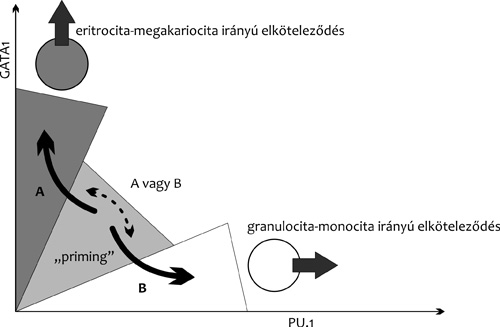
kifejeződése gátlás alá kerül. Ennek a keresztgátlásnak – aminek
pontos mechanizmusát ma sem ismerjük – az egyik legismertebb példája
a vérképző őssejtek eritrocita-megakariocita, illetve
granulocita-monocita irányú differenciálódásának szabályozása (5.
ábra). Az őssejtben mind a Gata1, mind a PU.1 gének
kifejeződnek. Ha a Gata1 TF-szintje emelkedik a sejtben, akkor a
PU.1 gén kifejeződése csökken, és a sejt hamarosan csak
eritrocita-megakariocita irányba képes differenciálódni. Fordított
esetben a Gata1 gén működése szűnik meg, és a sejt a
granulocita-monocita fejlődési irányba köteleződik el (Eguizabal et
al., 2013).
Végül: a legújabb transzdifferenciációs módszer a
„Jamanaka-faktorok” felhasználásán alapul. A sejtekbe – például
fibroblasztokba – bejuttatják az indukált pluripotens sejtek
előállításánál alkalmazott négy gént (Oct4, Sox2, Klf4, és c-Myc),
de nem várják meg a pluripotens sejtek kialakulását, hanem néhány
nap múlva egy morfogént (csont morfogenetikus fehéje-4-et) adnak a
tápfolyadékhoz. Ilyenkor a fibroblasztok idegsejtekké
transzdifferenciálódnak (Efe et al., 2011).
Néhány záró gondolat
A transzdifferenciáció, vagy, ahogy újabban gyakran nevezik:
fejlődési sorváltás (lineage conversion) tehát létező jelenség. Az
egyedfejlődés és a szövetregeneráció során in vivo is megfigyelhető,
in vitro pedig viszonylag egyszerűen kiváltható a „mester” TF-okat
kódoló gének manipulációjával. Ma már szinte hetente jelennek meg a
különböző szöveti őssejtek és véglegesen differenciálódott testi
sejtek sikeres újraprogramozásáról szóló beszámolók. Ez reményt
adhat olyan betegségek kezelésére is, amelyek rosszul vagy
egyáltalán nem osztódó testi sejtek (idegsejtek, inzulintermelő
béta-sejtek, porcsejtek) pusztulására vezethetők vissza. A módszer
nagy előnye lenne, hogy a pusztuló sejteket a beteg saját sejtjeivel
lehetne pótolni, így a beavatkozás után nem kellene immunológiai
komplikációkkal számolni. Megjósolni természetesen nem tudjuk,
hozhat-e áttörést a sejtek genetikai újraprogramozása az
orvostudományban, és ha igen, akkor ez mikor következik be. Annyit
azonban biztosan állíthatunk, hogy a transzdifferenciáció kutatása
már eddig is alapvetően megváltoztatta az egyedfejlődés,
differenciálódás és szövetregeneráció génszintű szabályozásáról
alkotott képüket.
Kulcsszavak: epigenetikai tájkép, imaginális korong, mester
transzkripciós faktor, szöveti őssejt, transzdifferenciáció
IRODALOM
Alison, M. R. – Poulsom, R. – Jeffery, R.
– Dhillon, A. P. – Quaglia, A. – Jacob, J. – Novelli, M. – Prentice,
G. – Williamson, J. – Wright, N. A. (2000): Hepatocytes from
Non-Hepatic Adult Stem Cells. Nature. 406, 257.
Davis, R. L. – Weintraub, H. – Lassar, A.
B. (1987): Expression of a Single Transfected cDNA Converts
Fibroblasts to Myoblasts. Cell. 51, 987–1000.
Day, R. C. – Beck, C. W. (2011):
Transdifferentiation from Cornea to Lens in Xenopus laevis Depends
on BMP Signalling and Involves Upregulation of Wnt Signalling. BMC
Developmental Biology. 11, 54.
Di Tullio, A. – Vu Manh, T. P. – Schubert,
A. – Castellano, G. – Månsson, R. – Graf, T. (2011): CCAAT/Enhancer
Binding Protein Alpha (C/EBP(Alpha))-Induced Transdifferentiation of
Pre-B Cells into Macrophages Involves No Overt Retrodifferentiation.
Proceedings of the National Academy of Sciences of the USA. 108,
17016–17021.
Efe, J. A. – Hilcove, S. – Kim, J. – Zhou,
H. – Ouyang, K. – Wang, G. – Chen, J. – Ding, S. (2011): Conversion
of Mouse Fibroblasts into Cardiomyocytes Using a Direct
Reprogramming Strategy. Nature Cell Biology. 13, 215–222.
Eguizabal, C. – Montserrat, N. – Veiga, A.
– Izpisua Belmonte, J. C. (2013): Dedifferentiation,
Transdifferentiation, and Reprogramming: Future Directions in
Regenerative Medicine. Seminars in Reproductive Medicine. 31, 82–94.
Gaustad, K. G. – Boquest, A. C. –
Anderson, B. E. – Gerdes, A. M. – Collas, P. (2004): Differentiation
of Human Adipose Tissue Stem Cells Using Extracts of Rat
Cardiomyocytes. Biochemical and Biophysical
Research Communications.; 314, 420–427.
Gussoni, E. – Soneoka, Y. – Strickland, C.
D. – Buzney, E. A. – Khan, M. K. – Flint, A. F. – Kunkel, L. M. –
Mulligan, R. C. (1999): Dystrophin Expression in the Mdx Mouse
Restored by Stem Cell Transplantation. Nature. 401, 390–394.
Halley, Julianne D. – Winkler, D. A. –
Burden, F. R. (2008): Toward a Rosetta Stone for the Stem Cell
Genome: Stochastic Gene Expression, Network Architecture, and
External Influences. Stem Cell Research. 1, 157–168.
Lagasse, E. – Connors, H. – Al-Dhalimy, M.
– Reitsma, M. – Dohse, M. – Osborne, L. – Wang, X. – Finegold, M. –
Weissman, I. L. – Grompe, M. (2000): Purified Hematopoietic Stem
Cells Can Differentiate into Hepatocytes in vivo. Nature Medicine.
6, 1229–1234.
Laiosa, C. V. – Stadtfeld, M. – Xie, H. –
De Andres-Aguayo, L. – Graf, T. (2006): Reprogramming of Committed T
Cell Progenitors to Macrophages and Dendritic Cells by C/EBP Alpha
and PU. 1 Transcription Factors. Immunity. 25, 731–744.
Massengale, M. – Wagers, A. J. – Vogel, H.
– Weissman, I. L. (2005): Hematopoietic Cells Maintain Hematopoietic
Fates upon Entering the Brain. The Journal of Experimental Medicine.
201, 1579–1589.
Maves, L. – Schubiger, G. (1999): Cell
Determination and Transdetermination in Drosophila Imaginal Discs.
Current Topics in Developmental Biology. 43, 115–151.
Nutt, S. L. – Heavey, B. – Rolink, A. G. –
Busslinger, M. (1999): Commitment to the B-Lymphoid Lineage Depends
on the Transcription Factor Pax5. Nature. 401, 556–562.
Orlic, D. – Kajstura, J. – Chimenti S. –
Jakoniuk, I. – Anderson, S. M. – Li, B. – Pickel, J. – Mckay, R. –
Nadal-Ginard, B. – Bodine, D. M. – Leri, A. – Anversa, P. (2001):
Bone Marrow Cells Regenerate Infarcted Myocardium. Nature. 410,
701–705.
Park, K. S. – Wells, J. M. – Zorn, A. M. –
Wert, S. E. – Laubach, V. E. – Fernandez, L. G. – Whitsett, J. A.
(2006): Transdifferentiation of Ciliated Cells During Repair of the
Respiratory Epithelium. American Journal of Respiratory Cell and
Molecular Biology. 34, 151–157.
Patapoutian, A. – Wold, B. J. – Wagner, R.
A. (1995): Evidence for Developmentally Programmed
Transdifferentiation in Mouse Esophageal Muscle. Science. 270,
1818–1821.
Perán, M. – Marchal, J. A. –
Rodríguez-Serrano, F. – Alvarez, P. – Aránega, A. (2011):
Transdifferentiation: Why and How? Cell Biology International. 35,
373–379.
Takahashi, K. (2012): Cellular
Reprogramming—Lowering Gravity on Waddington’s Epigenetic Landscape.
Journal of Cell Science. 125, 2553–2560.
Theise N. D. (2010): Stem Cell Plasticity:
Recapping the Decade, Mapping the Future. Experimantal Hematology.
38, 529–539.
Vassilopoulos, G. – Wang, P. R. – Russell,
D. W. (2003): Transplanted Bone Marrow Regenerates Liver by Cell
Fusion. Nature. 422, 901–904.
Vierbuchen, T. – Ostermeier, A. – Pang, Z.
P. – Kokubu, Y. – Südhof, T. C. – Wernig, M. (2010): Direct
Conversion of Fibroblasts to Functional Neurons by Defined Factors.
Nature. 463, 1035–1041.
Waddington, Conrad Hal (1957): The
Strategy of the Genes. Allen & Unwin, London
Wagers, A. J. – Weissman, I. L. (2004):
Plasticity of Adult Stem Cells. Cell. 116, 639–648.
Yang, L. – Li, S. – Hatch, H. – Ahrens, K.
– Cornelius, J. G. – Petersen, B. E. – Peck, A. B. (2002): In Vitro
Trans-differentiation of Adult Hepatic Stem Cells into Pancreatic
Endocrine Hormone-producing Cells. Proceedings of the National
Academy of Sciences of the USA. 99, 8078–8083.
Yoon, J. – Shim, W. J. – Ro, Y. M. – Lim,
D. S. (2005): Transdifferentiation of Mesenchymal Stem Cells into
Cardiomyocytes by Direct Cell-to-Cell Contact with Neonatal
Cardiomyocyte But Not Adult Cardiomyocytes. Annals of Hematology.
84, 715–721.
|