|
|
Az embrionális, illetve az indukált pluripotens
őssejtek szinte korlátlan fejlődési képességgel rendelkező
sejtformák. Nem véletlen, hogy többféle betegség sejtterápián
alapuló gyógyítását e sejtekből kiindulva próbálják kidolgozni. E
közlemény röviden ismerteti a pluripotens őssejtek legfontosabb
tulajdonságait, és megpróbál választ adni arra, mi teszi ezeket a
sejteket oly különlegessé és plasztikussá.
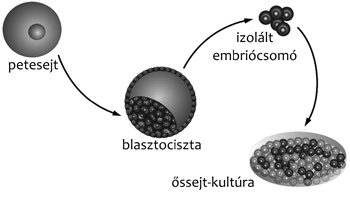
Az embrionális őssejtek eredete és létrehozása
Ahhoz, hogy a pluripotens őssejtek biológiai programját megismerjük,
fontos felvázolni, hogy e sejtek honnan erednek, milyen forrásból
hozhatók létre. A többsejtű élőlények egyetlen sejt, a
megtermékenyített petesejt osztódása révén indulnak fejlődésnek. A
megtermékenyített petesejt (zigóta) a kiindulási alapja a későbbi
élőlénynek, tehát olyan totipotens alapítósejt, amiből az egész
szervezet létrejön. Emlősöknél ebből alakul ki az embrió, továbbá e
sejt leszármazottai hozzák létre az extraembrionális képletek, így a
méhlepény magzati eredetű részét is. A zigóta, fejlődése során több
átalakuláson megy keresztül, osztódás révén elsőként a néhány
sejtből álló szedercsíra (blasztula) jön létre. A blasztula állapot
korai fázisában (négysejtes állapot) az individuális sejtek még
megőrzik totipotens jellegüket, egymástól elkülönítve is képesek
létrehozni az egész élőlényt. Ehhez hasonló természetes folyamat
vezethet a többpetéjű ikrek kialakulásához.
Az embriógenezis következő fázisában, a hólyagcsíra
(blasztociszta) állapotban a sejtek elvesztik totipotens jellegüket,
azaz az individuális sejtekből már nem képes kialakulni a teljes
szervezet, viszont az egyes sejtek még mindig széles fejlődési
potenciállal rendelkeznek. Korai blasztociszta állapotban (egérnél
3,5 nappal a megtermékenyítés után) az embriókezdemény körülbelül
húsz-negyven sejtből áll. A külső sejtréteget trofektodermának
nevezzük; e sejtek nem vesznek részt a későbbi magzat
kialakításában, viszont ebből alakul ki a méhlepény nem anyai
eredetű rétege. A blasztociszta belsejében található az úgynevezett
belső sejttömeg (inner cell mass), amit embriócsomónak is neveznek.
A korai blasztociszta állapotban az egér embriócsomó többé-kevésbé
homogén sejtcsoportnak tekinthető. Ezzel szemben az embrió
implantációja során, tehát a késői blasztociszta állapotban (egérnél
4,5 nappal a megtermékenyítés után), az embriócsomó sejtjei már
egyértelműen elkülönülnek epiblaszt és hipoblaszt sejtekre. A belül
elhelyezkedő epiblaszt sejtekből jön létre az embrió összes sejt- és
szövetfélesége. A hipoblaszt sejtek az extraembrionális szikhólyag
endodermális részének kialakításában vesznek részt, a későbbi embrió
felépítésében nem. Az embriógenezis következő fázisában
(gasztruláció) az embriócsomó elkülönült epiblaszt sejtjei egyrétegű
struktúrát hoznak létre, majd e sejtréteg szakaszosan átalakul
ektodermális, endodermális és mezodermális sejtréteggé. A fejlődés
ezen fázisában kezdenek specializálódni a sejtek, ezekből az
átalakult sejtformákból jönnek létre a különféle szervkezdemények.
Blasztociszta állapotban tehát elkülönülnek azok a
sejtek (az embriócsomó sejtjei, majd ezekből az epiblaszt sejtek),
amelyekből a későbbi embrió képződik. Ezek a sejtek már nem
totipotens, hanem pluripotens tulajdonsággal rendelkeznek. A
pluripotens jelleg azt jelenti, hogy még mindig széles fejlődési
potenciállal bírnak, de mindent már nem képesek létrehozni. A
pluripotens sejtekre az jellemző, hogy az extraembrionális sejtek
egy részének kivételével, az összes testi sejt kialakulhat belőlük.
Fontos kiemelni, hogy az embriócsomóban létrejött pluripotens
őssejtek a fejlődő szervezetben csak átmenetileg léteznek, a
differenciálódás folyamán fokozatosan alakulnak át specializáltabb
és szűkebb fejlődési potenciállal rendelkező sejtekké. Mesterséges
környezetben, az embrióból kiszakítva, megfelelő körülmények között
viszont e pluripotens őssejtek korlátlanul fenntarthatók, s megőrzik
az alapító sejtekre jellemző tulajdonságaikat.
Az embrionális őssejtek (embryonic stem – ES) tehát
az epiblaszt sejtek előalakjaiból, az embriócsomó sejtjeiből
hozhatók létre. Egér ES-sejteket elsőként Martin Evans és Matthew
Kaufman kutatóknak, illetve ezzel párhuzamosan Gail Martinnak
sikerült létrehoznia több mint harminc évvel ezelőtt (Evans, 2011).
Evans és Kaufman az ES-sejteket a blasztociszta letapasztása után,
az embriócsomóból kinövő sejtekből hozta létre, míg Gail Martin
izolálta a belső embriócsomót, és ezekből sikerült ES-sejteket
felszaporítania. A későbbiekben az egyszerűsége miatt a
blasztociszta letapasztásos módszer terjedt el, de ennél az
eljárásnál is az embriócsomóból származnak az ES-sejtek.
Hozzávetőleg húsz évvel később, 1998-ban sikerült James Thomson
munkacsoportjának létrehoznia az első humán ES-sejteket, emberi
petesejtek mesterséges megtermékenyítése során kapott blasztociszták
felhasználásával (Thomson et al., 1998). A humán ES-sejtek
létrehozása nagy publicitást kapott, sok kutató hatalmas áttörésnek
tekintette ezt a felfedezést, mert itt már nem kísérleti állatokból
származó sejtekről volt szó, hanem olyan emberi sejtforrásról, amit
a regeneratív medicina fel tud használni. Ezzel párhuzamosan
megindultak az etikai/jogi viták és korlátozások, mivel e sejtek
létrehozása során a fejlődő blasztocisztát fel kellett áldozni az
ES-sejtek kinyerése céljából.
A múlt század végén tehát megszülettek azok a
technológiák, amelyekkel pluripotens őssejtek váltak létrehozhatóvá
embriókezdeményből. A területen a következő nagy áttörést Jamanaka
Sinja (Shinya Yamanaka) és Takahasi Kazutosi (Kazutoshi Takahashi)
felfedezése jelentette, akiknek embrió felhasználása nélkül, felnőtt
sejtek visszaprogramozásával sikerült előállítaniuk úgynevezett
indukált pluripotens őssejteket (iPS-sejteket) (Takahashi –
Yamanaka, 2006). Ez a felfedezés külön cikkben kerül ismertetésre, e
közleményben inkább az ES-/iPS-sejtek különleges sajátságait
szeretnénk bemutatni és értelmezni.
A pluripotens őssejtek jellegzetességei
Az iPS-, illetve az ES-sejtek legértékesebb sajátsága a
pluripotencia, azaz, hogy belőlük a legkülönfélébb sejtformák
fejlődhetnek ki. Az átalakulási képesség mellett az ilyen sejtek
megfelelő kondíciók esetén korlátlan önmegújuló sajátsággal is
rendelkeznek. Ezt a jellegüket nagyszámú sejtosztódást követően is
képesek megőrizni. Mi teszi a pluripotens őssejteket ilyen
különlegessé; ellentétben a szöveti őssejtekkel, miért tudnak szinte
minden irányba átalakulni? Erre az egyszerű válasz, hogy az őssejtek
totipotens-pluripotens jellege egy szükséges tulajdonság, mivel az
összes sejtünk egyetlen forrásból, a megtermékenyített petesejtből
származik, amelynek közvetlenül vagy közvetetten, de minden irányba
tudnia kell az átalakulást. Korábban felvázoltuk, hogy a totipotens,
majd a pluripotens jelleget fokozatosan vesztik el az
embriókezdeményt alkotó sejtek. Úgy tűnik, az embrionális
őssejtforma olyan alapállapot, amelybe vissza lehet programozni a
sejteket, illetve megfelelő szignálok hatására e metastabil állapot
fenn is tartható. A következőkben ismertetésre kerül, hogy milyen
faktorok, illetve szignálok képesek ezt a pluripotens állapotot
fenntartani, stabilizálni.
Embrionális őssejtekben olyan kulcsgéneket
azonosítottak, melyek esszenciálisak a pluripotens állapot
eléréséhez és stabilizálásához. Ezek a pluripotencia faktorok a
transzkripciós faktorok családjába tartoznak, melyek a gének
átírását (transzkripcióját) modulálják. Pluripotens állapot
fenntartásában számos transzkripciós faktor vesz részt, de ezek
közül három fehérjének az Oct4, a Sox2 és a Nanog nevű
transzkripciós faktoroknak van kitüntetett szerepük. E kulcs
transzkripciós faktorok további transzkripciósfaktor-hálózatokat
aktiválnak, melyek elősegítik a differenciálatlan állapot
fenntartását. Továbbá ezen három kulcsfehérje számos, a sejtek
továbbalakulását elősegítő gént gátol. Érdekes módon nem teljes
gátlásról van szó, inkább e sejtekre az jellemző, hogy génjeik egy
része bivalens állapotban van, azaz készek az átalakulásra, csak egy
jelre várnak (Bernstein et al., 2006). Ezt az állapotot a
DNS-fehérjekomplex (kromatin) egy jellegzetes módosult formájával
lehet jellemezni, ami specifikus az embrionális őssejtekre. Fontos
még egyszer kihangsúlyozni, hogy a pluripotens állapot az
egyedfejlődés során átmeneti jellegű, a sejtek spontán próbálnak
ebből az állapotból kilépni, és elindulni egyik vagy másik fejlődési
irányba. Mindezek miatt az embrionális őssejtek metastabil
állapotban vannak, ami csak mesterséges körülmények között tartható
fenn, megfelelő szignálok segítségével. E szolubilis és sejtfelszíni
molekulák által közvetített szignálok természete az utóbbi
évtizedben vált ismertté.
Az ES-sejtek mesterséges körülmények közötti
fenntartásának optimalizálása hosszú folyamat volt, mely során
fokozatosan sikerült azonosítani azokat a kulcsszignálokat, amelyek
biztosítják a pluripotens állapot stabilitását. ES-sejteket
kezdetben embrionális dajkasejtekkel (egér embrionális kötőszöveti
sejtekkel) tenyésztettek együtt (Thomson et al., 1998). A
dajkasejtek képesek voltak fenntartani az ES-sejtek pluripotens
jellegét, amire többek között a sejtek morfológiai és funkcionális
tulajdonságai alapján lehetett következtetni. Későbbiekben
azonosították azokat a szolubilis faktorokat, melyeket a dajkasejtek
termeltek, s kulcsfontosságúak az ES-sejtállapot megőrzéséhez.
Érdekes módon a humán és az egér ES-sejtek tenyésztéséhez nem
ugyanazok a faktorok szükségesek, ami elég meglepő, ha
feltételezzük, hogy a korai embriógenezis hasonló lépések
sorozatából áll. Egér őssejteket dajkasejtek nélkül, LIF (leukemia
inhibitory factor) jelenlétében lehet fenntartani (Ying et al.,
2008), humán ES-sejtek tenyésztéséhez viszont bFGF- (basic
fibroblast growth factor) faktorra van szükség. Újabb vizsgálatok
szerint egérből is lehet olyan pluripotens sejteket izolálni, melyek
bFGF-t igényelnek (Nichols – Smith, 2009). Ezek a sejtek már
fejlettebb állapotban lévő, implantáció utáni egérembrió epiblaszt
sejtjeiből nyerhetők ki, ezért ezeket epiblaszt őssejteknek nevezik.
Az egérből származó epiblaszt őssejtek morfológiája is hasonló a
humán ES-sejtekhez: ellentétben a dóm formájú kolóniákat képező egér
ES-sejtekkel, ezek a sejtek laposabb formájú telepeket képeznek, és
szintén érzékenyek a tripszines emésztésre.
Ezen eredmények alapján kialakult a koncepció, mely
szerint a pluripotens sejteknek legalább kétféle formája létezik. Az
egér embriócsomóból izolált sejteket nevezik naiv ES-sejteknek, ezek
reprezentálják az embrionális sejtek alapállapotát, melyek teljes
értékű őssejteknek tekinthetők. A későbbi fázisban izolálható
„aktivált” (primed) őssejtek reprezentálják azt a sejtpopulációt,
amely már előrehaladottabb, úgymond készen áll a differenciálódásra
(Nichols – Smith, 2009). A humán ES-sejtek – annak ellenére, hogy a
korai embriócsomóból származnak – sokkal jobban hasonlítanak az egér
epiblaszt őssejtekhez, ami arra utal, hogy ezek már
elkötelezettebbek. A felvázolt különbségek ellenére az emlős
pluripotens őssejtek több közös morfológiai és funkcionális
tulajdonsággal rendelkeznek. A korábban már említett három
pluripotencia faktor kimutatható szinte minden pluripotens
őssejtben, ezért a morfológiai jellegek mellett e faktorok
pozitivitása alapján szokás értékelni az embrionálisőssejt-kolóniák
minőségét. Összefoglalva, a pluripotens őssejtek olyan átmeneti
állapotban vannak, melyek még nyitottak minden fejlődési irányra,
csak egy külső jelre várnak, hogy beinduljon egy differenciálódási
program. Persze önmagában a pluripotens alapállapot fenntartása túl
sok gyakorlati haszonnal nem kecsegtet, e sejtek képességeit úgy
lehet kiaknázni, ha átalakítjuk őket.
A pluripotens őssejtek
in vivo és in vitro átalakulási képessége
Az egér ES-/iPS-sejtek pluripotens jellege azt is jelenti, hogy
korai embriókezdeménybe visszajutatva őket, néhány extraembrionális
képlet kivételével, képesek az egész szervezet létrehozására. Az ES-
vagy iPS-sejtek, tehát képesek beintegrálódni az embriókezdeménybe,
és ugyanazt a fejlődési programot hajtják végre, mint a barázdálódás
révén keletkező embriócsomó sejtjei. Ez a fejlődési program több
intermedier állapoton keresztül valósul meg, a sejtek átalakulását
hormonszerű szolubilis és sejtfelszíni molekulák irányítják, melyek
kulcsgének be- és kikapcsolásával szabályozzák a fejlődést.
Nyilvánvaló, hogy ilyen jellegű in vivo vizsgálatok
csak állatokon végezhetők el, etikai, illetve praktikus szempontok
miatt ilyen teszteket emberi ES-sejtekkel, humán embriókon nem lehet
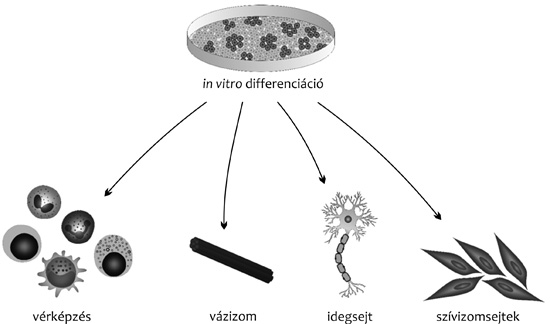
végrehajtani. Humán mintáknál az ES-sejtek in vitro
differenciálására törekszünk, ami a kutatásban és a klinikumban is
hasznosulhat. Fontos célkitűzés és egyben nagy kihívás funkcionális
formákká alakítani át az őssejtállapotból e sejteket, melyek
kísérleti vagy esetleg terápiás célra is felhasználhatók. A
pluripotens sejtek legfőbb előnye a szöveti multipotens őssejtekhez
képest, hogy belőlük a szervezetet alkotó összes sejttípus
kifejlődhet. Így elvileg segítségükkel minden hiányzó vagy
működésképtelen sejtet újra lehet gyártani. A pluripotens
őssejtekből létre lehet hozni többek között az ideg-, az emésztő-,
az ér- és a vázizomrendszer sejtjeit és szöveteit.
Annak ellenére, hogy egér ES-sejtek in vivo
tökéletesen át tudnak alakulni az embrió összes sejttípusává,
mesterséges körülmények között még mindig nagy kihívás hatékonyan
irányítani a sejtek fejlődését. A legtöbb sejtátalakítási protokoll
az in vivo folyamatokat próbálja utánozni, mesterségesen biztosítva
azokat a faktorokat, melyek az embriógenezis során termelődnek
(Keller, 2005).
|
|
|
Ugyanakkor az igazsághoz hozzátartozik, hogy a
kutatók sok esetben próbálgatás útján dolgozták ki azokat a
differenciálási eljárásokat, melyekkel egy-egy specifikus sejttípus
létrejön. Hasonlóan a normál embrionális fejlődéshez, a pluripotens
sejtek mesterséges átalakulása is hosszadalmas folyamat része: az
őssejtek elsőként multipotens sejtekké alakulnak át, majd ezek
formálódnak tovább egy vagy több lépésen keresztül terminálisan
differenciálódott sejtekké. Mind az embrionális fejlődés során, mind
az őssejtek mesterséges átalakításakor nehéz definiálni, mikor
veszíti el a sejt pluripotens jellegét, illetve mely pillanattól
köteleződik el az adott fejlődési irányba. Az ES-sejtek átalakítása
alapvetően háromféle technikával valósítható meg (Keller, 2005).
ES-sejtek összetapadásával háromdimenziós aggregátumok, úgynevezett
embrioid testek képződhetnek, melyek képesek növekedni, és az őket
alkotó sejtek képesek fokozatosan átalakulni. A másik lehetőség
tápláló dajkasejteken növeszteni az őssejteket, ekkor a dajkasejtek
modulálják a sejtfejlődést különféle faktorok termelése révén.
Például az OP9 dajkasejtek elősegítik a vérképző sejtek létrejöttét.
A harmadik módszer a sejtek fehérjemátrix felületen való
tenyésztésén alapul; ennél a megközelítésnél hormonok, illetve
növekedési faktorok adagolásával indul el a sejtek átalakulása.
Mindhárom módszernek megvannak az előnyei és hátrányai. Az embrioid
testek előnye, hogy a létrejövő háromdimenziós sejtaggregátum
emlékeztet a korai embriókezdeményre, a különféle sejtek kölcsönösen
indukálják egymás fejlődését. A módszer hátránya, hogy a folyamatot
nehéz kontrollálni, mivel az embrioid testekben nemcsak a létrehozni
kívánt sejttípusok keletkeznek, hanem szinte mindenféle sejtféleség,
s ezek aránya is különböző lehet. A dajkasejtek felhasználásával
történő sejtátalakítás irányítottabb sejtfejlődést eredményez, mivel
a dajkasejtek olyan faktorokat termelnek, melyek egy adott sejttípus
fejlődését és túlélését segítik elő. Ennél a megközelítésnél
hátrányt jelent, hogy a sejtek átalakítását követően a
sejtkultúrában ottmaradnak a dajkasejtek is, melyektől sokszor nehéz
elválasztani az ES-eredetű differenciált sejteket. A harmadik
módszernél tudjuk legjobban kontrollálni a sejtek fejlődését,
ugyanakkor a proteinmátrix természetétől függően változhat a sejtek
átalakulási és túlélési képessége. Ezek az eljárások az ES-sejtek
kezdeti átalakítására alkalmasak, az így létrejövő sejtek általában
multipotens alakok, melyeket megfelelő faktorokkal tovább lehet
alakítani adott sejttípussá.
Vérképző sejtek létrehozása
pluripotens őssejtekből
Ezen általános összefoglaló munka kereteit meghaladja a különféle
irányú ES-sejt-átalakítások ismertetése, ehelyett egyetlen irány, a
vérképző sejtek létrehozása kerül bemutatásra. Ha pluripotens őssejt
eredetű hemopoetikus (vérképző) sejteket akarunk létrehozni, akkor
kétféle célkitűzésről lehet szó. Az ambiciózusabb cél a teljes
értékű vérképző őssejtek (hematopoietic stem cell – HSC)
létrehozása. A másik lehetőség érett sejtek előállítása, például
vörösvérsejtek, vérlemezkék vagy fehérvérsejtek gyártása. A HSC-k
direkt előállításának előnye, hogy ezeket visszaültetve a
szervezetbe korlátlan ideig biztosítani tudják a vérképzést.
Ugyanakkor még ma is hézagosak az ismereteink a HSC-k eredetéről,
ami nehézzé teszi az embrionális őssejtekből történő létrehozásukat.
A korai embriogenezis során a hemopoetikus sejtek elsőként az
extraembrionális szikzacskóban alakulnak ki, majd az embrionális
aorta falában, illetve az extraembrionális artériákban is beindul a
vérképző sejtek fejlődése. Az embrionális fejlődés későbbi
szakaszában a vérképzés fokozatosan áttevődik a magzati májba, majd
a születést követően végső helye a vörös csontvelő lesz. A képet
tovább bonyolítja, hogy nem azonos a különféle anatómiai területeken
létrejövő vérképző sejtek fejlődési képessége. A ma leginkább
elfogadott elmélet szerint nem az extraembrionális szikzacskóban,
hanem az embrió belsejében, a fejlődő aorta falában jönnek létre a
HSC-k előalakjai (Orkin – Zon, 2008). Ezeket az ismereteket is
figyelembe véve számos kutatócsoport próbált és próbál vérképző
őssejtet létrehozni ES-, illetve iPS-sejtekből. A HSC-sajátság
igazolására a leginkább elfogadott módszer a transzplantációs
vizsgálat, mely teszteli, hogy a létrehozott vérképző progenitorok a
kísérleti állatokba beültetve mennyi ideig, illetve milyen típusú
sejteket képesek termelni. Számos ES-/iPS-sejtből származó
HSC-transzplantációs vizsgálat történt, ugyanakkor senkinek sem
sikerült reprodukálhatóan és genetikai módosítás nélkül olyan
hemopoetikus sejtet létrehoznia, mely hosszú távon fenntartotta
volna a vérképzést. Ugyanakkor magzatból ki lehet nyerni olyan
HSC-ket, melyek képesek erre, tehát már embrionális korban
megjelennek az ilyen tulajdonságokkal rendelkező sejtek. Hogy miért
nem tudjuk reprodukálni mesterséges körülmények között az in vivo
bekövetkező HSC-képződést, annak több oka is lehet. Egyrészt, ahogy
az előzőekben felvázoltuk, nem teljesen világos, hogy az első HSC-k
in vivo milyen átmeneti formákból jönnek létre, illetve az is
tapasztalati tény, hogy a felnőtt állatból izolált HSC-k in vitro
nagyon hamar elvesztik az őssejt-tulajdonságukat. A HSC-k
előállításának sikertelensége egy fontos tényre hívja fel a
figyelmet: az elméletileg mindenre képes embrionális őssejteket
mesterséges körülmények között ma még nem tudjuk akármivé
átalakítani.
A vérképző őssejtek előállítása mellett a másik
lehetőség az érett, kész vérsejtek létrehozása pluripotens
őssejtekből. Ez az eljárás hatékonyabbnak bizonyult, mivel
vörösvértesteket, vérlemezkéket, illetve különféle fehérvérsejteket
sikerült generálni pluripotens őssejtekből (Olsen et al., 2006).
Ugyanakkor ezek az átalakítási kísérletek egy másik fontos
aspektusra hívják fel a figyelmet. Ismert, hogy a fejlődés során a
vörösvértestekben található oxigénkötő fehérje, a hemoglobin más
változata fordul elő a korai embrionális, a magzati, illetve a
születés utáni korban. Megkülönböztetünk embrionális és magzati
hemoglobinokat, melyek tulajdonságai eltérőek a felnőttekben lévő
fehérjékhez képest. Az embrionális, illetve a magzati hemoglobinok
nagyobb affinitással kötik az oxigént, ami elősegíti az oxigén
átjutását az anyai vérből a magzati vérbe. Érdekes módon a humán
ES-sejtekből létrehozott vörösvértestek főleg embrionális és magzati
hemoglobinokat termelnek, de a felnőttekre jellemző formák csak kis
mennyiségben detektálhatóak (Chang et al., 2006). Ez persze nem
meglepő, hiszen a normál embriógenezis során is ezek a fehérjeformák
jelennek meg elsőként a szikzacskóban kialakuló vérképzés során, s
valószínűleg az ES-sejtekben is ezek a fejlődési programok tudnak
legkönnyebben aktiválódni. Feltételezhető, hogy ez a jelenség nem
korlátozódik a vörösvértestekre, valószínű más sejteknél is az
embrionális fejlődési programok aktiválódnak leghatékonyabban. Ez a
megfigyelés rávilágít az ES-/iPS-eredetű sejtek átalakításának egy
másik korlátjára. Valószínű, hogy a pluripotens sejtekből
létrehozott funkcionális formáknak nincsenek a felnőtt sejtekre
jellemző tulajdonságaik, ezért visszaültetve a szervezetbe nem
biztos, hogy ugyanolyan hatékonyan működnek, mint a felnőtt
szervezetből származó sejtek. Mindezek a megfigyelések felhívják a
figyelmet arra, hogy mesterséges körülmények között, az
ES-/iPS-sejtek fejlődési potenciáljának kiaknázása céljából további
intenzív kutatásokra van szükség.
A pluripotens őssejt felhasználásának korlátai
Az eddigiekben a pluripotens őssejteket úgy jellemeztük, hogy
képesek önmegújulásra, illetve több-kevesebb megszorítással képesek
mindenféle sejtformává átalakulni. Felmerülhet a kérdés, hogy ha
ilyen kedvező sajátságaik vannak, akkor miért nem használják őket
tömegesen a sejtterápiás eljárások során. A pluripotens jellegnek
veszélyes következménye is van, ugyanis ha visszainjektáljuk az
ilyen sejteket a recipiens szervezetbe, akkor azokban speciális
tumorok, teratómák fejlődnek ki. A teratómák jóindulatú daganatok,
melyek az őssejtből spontán differenciálódott sejtek tömegéből
állnak. A pluripotens őssejtekre jellemző, hogy mindhárom csíralemez
(ekto-, endo- és mezoderma) sejtjeit/szöveteit tartalmazó tumorokat
képesek létrehozni. Leegyszerűsítve: a pluripotencia és a
tumorgenezis együttesen jellemző az ES-/iPS-sejtekre.
Szerencsére a pluripotens sejtekből létrejövő
differenciálódott formák fokozatosan elveszítik a tumorképző
sajátságukat, de ez nagyban függ a sejtátalakítás hatékonyságától,
illetve az átmeneti formák tisztaságától. Például
vázizom-differenciálás során azt tapasztalták, hogy ötnapos
tenyésztéssel embrioid testekké differenciált, majd további 7–10
napig tenyésztett egér embrionális őssejtből származó izom
progenitorokat visszajuttatva immunhiányos egerekbe, tumorok
alakultak ki. Ám ha az öt napig differenciáltatott sejtek közül
szortolással (áramlási citometriával) kiválogatták és tovább
növesztették az izomsejt-előalakokat, akkor az állatokba injektált
sejtekből nem jött létre teratóma, hanem csak izomsejtek fejlődtek
ki (Darabi et al., 2008). Feltételezhető, hogy sejtválogatás nélkül,
a több mint tíz napig tartó sejtdifferenciálás ellenére maradtak a
sejttenyészetben formák, melyek megőrizték daganatképző embrionális
őssejt jellegüket. Humán ES-sejteknél is megfigyelték, hogy tizenhat
napig tartó differenciáltatást követően patkányokba transzplantált
idegsejt progenitorokból tumorok fejlődtek. Igaz, a hosszabb ideig
(huszonkét napig) differenciáltatott sejtekből rákos sejtburjánzás
már volt detektálható (Brederlau et al., 2006). Ezek az eredmények
felhívják a figyelmet, hogy az ES-sejtek differenciálódásának
hosszától és hatékonyságától is függ, hogy a sejteknek megmarad-e
vagy sem a daganatképző sajátosságuk. Persze adódik a kérdés, hol a
határ, mikortól lehetünk teljesen biztosak abban, hogy a terápiás
célból használni kívánt ES-eredetű sejtkoktél nem tartalmaz
daganatképző sejteket. Ezt nagyon nehéz definiálni, mindezek miatt a
daganatos sejtburjánzás veszélye a legnagyobb korlátja a pluripotens
őssejtek terápiás használatának.
A sejtterápiás beavatkozások biztonsági kockázata
ellenére a pluripotens őssejtek ma már nélkülözhetetlenek a modern
sejtbiológiai és molekuláris medicina kutatásaihoz. A pluripotens
őssejtek használata már napjainkban is széles körben elterjedt,
feltételezhetően a jövőben mind több és több kutató – és klinikai –
laboratórium fog ilyen sejteket tenyészteni. Ugyanakkor még további
ismeretekre kell szert tennünk, hogy kontrolálni tudjuk e sejtek
átalakítását, ami lehetővé teszi a különféle altípusú és aktivitású
differenciál sejtek előállítását őssejtekből. Ehhez nagy segítséget
fognak nyújtani a kifejlesztett szintetikus tápfolyadékok és
adalékok, melyek lehetővé teszik, hogy a különböző laboratóriumokban
egységes protokollok segítségével tudják létrehozni és átalakítani e
sejteket. A standardizált procedúráknak különösen a klinikai
sejtterápiás vizsgálatoknál lesz fontos szerepük, hogy minden beteg
hasonló tulajdonságú biológiai mintával legyen kezelve.
Köszönet jár Sári Erikának az ábrák elkészítéséért. A szerző
munkáját a Bolyai János Kutatási ösztöndíj támogatása segítette.
Kulcsszavak: Embrionális őssejtek, pluripotens őssejtek,
transzkripciós faktorok, irányított sejtdifferenciálás,
embriógenezis
IRODALOM
Bernstein, B. E. – Mikkelsen, T. S. – Xie,
X. – Kamal, M. – Huebert, D. J. – Cuff, J. – Fry, B. – Meissner, A.
– Wernig, M. – Plath, K. et al. (2006): A Bivalent Chromatin
Structure Marks Key Developmental Genes in Embryonic Stem Cells.
Cell. 125, 315–326.
Brederlau, A. – Correia, A. S. – Anisimov,
S. V. – Elmi, M. – Paul, G. – Roybon, L. – Morizane, A. – Bergquist,
F. – Riebe, I. – Nannmark, U. et al. (2006): Transplantation of
Human Embryonic Stem Cell-derived Cells to a Rat Model of
Parkinson’s Disease: Effect of In Vitro Differentiation on Graft
Survival and Teratoma Formation. Stem Cells. 24, 1433–1440.
Chang, K. H. – Nelson, A. M. – Cao, H. –
Wang, L. – Nakamoto, B. – Ware, C. B. – Papayannopoulou, T. (2006):
Definitive-like Erythroid Cells Derived from Human Embryonic Stem
Cells Coexpress High Levels of Embryonic and Fetal Globins with
Little or No Adult Globin. Blood. 108, 1515–1523.
Darabi, R. – Gehlbach, K. – Bachoo, R. M.
– Kamath, S. – Osawa, M. – Kamm, K. E. – Kyba, M. – Perlingeiro, R.
C. (2008): Functional Skeletal Muscle Regeneration from
Differentiating Embryonic Stem Cells. Nature Medicine. 14, 134–143.
Evans, M. (2011): Discovering
Pluripotency: 30 Years of Mouse Embryonic Stem Cells. Nature Reviews
Molecular Cell Biology. 12, 680–686.
Keller, G. (2005): Embryonic Stem Cell
Differentiation: Emergence of a New Era in Biology and Medicine.
Genes & Development. 19, 1129–1155.
Nichols, J. – Smith, A. (2009): Naive and
Primed Pluripotent States. Cell Stem Cell. 4, 487–492.
Olsen, A. L. – Stachura, D. L. – Weiss, M.
J. (2006): Designer Blood: Creating Hematopoietic Lineages from
Embryonic Stem Cells. Blood. 107, 1265–1275.
Orkin, S. H. – Zon, L. I. (2008):
Hematopoiesis: An Evolving Paradigm for Stem Cell Biology. Cell.
132, 631–644.
Takahashi, K. – Yamanaka, S. (2006):
Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult
Fibroblast Cultures by Defined Factors. Cell. 126, 663–676.
Thomson, J. A. – Itskovitz-Eldor, J. –
Shapiro, S. S. – Waknitz, M. A. – Swiergiel, J. J. – Marshall, V. S.
– Jones, J. M. (1998): Embryonic Stem Cell Lines Derived from Human
Blastocysts. Science. 282, 1145–1147.
Ying, Q. L. – Wray, J. – Nichols, J. –
Batlle-Morera, L. – Doble, B. – Woodgett, J. – Cohen, P. – Smith, A.
(2008): The Ground State of Embryonic Stem Cell Self-renewal.
Nature. 453, 519–523.
|
|