|
Elsőként a lúdfű (Arabidopsis thaliana), majd a
rizs (Oriza sativa) szekvencia-adatbázisait közölték. A szekvenátorok
új generációinak kifejlesztése révén ma már egyre több gazdaságilag
fontos növényünk DNS-szekvenciája ismert (lásd: CoGePedia). A teljes
genomszekvencia-adatok mellett fontos információt hordoznak a rövid
genomszakaszok adatai (Genome Survey Sequencing – GSS), illetve a
kifejeződő géneket reprezentáló, az RNS-molekulákról készített kópia
cDNS-ek szekvenciáinak adatbázisai. A plantgdb.org weboldalon
megtalálhatók ezek az adatok.
A genomszekvenálási projektek jelentősége abban is
megmutatkozik, hogy az agronómiai szempontból fontos gének
azonosításának és izolálásának lehetőségei nagymértékben
kiszélesedtek. A polimeráz láncreakció (PCR) módszer segítségével még
a kis példányszámú, specifikus DNS-szekvenciarészek is megsokszorozva
megszintetizálhatók, és így a gének izolálhatók (Dudits, 2009). Az
érdekes gének keresésében fontos támpontot nyújtanak a génkifejeződési
mintázatok, amelyeket Gén-Csip technológiával akár a genom egészére
vonatkozóan is meg lehet állapítani. A növényi gének és a
növény-patogén szervezetek génjeinek kifejeződési mintázatairól
adatbázisokban tájékozódhatunk (PLEXdb [URL1]). A gének aktivitása
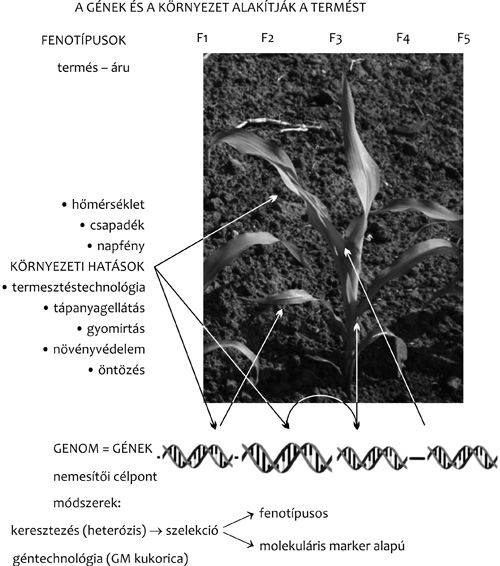
igen eltérő lehet a különböző szervekben, szövetekben, és a környezeti
tényezők hatásaira adott élettani válaszok elsődlegesen a
génkifejeződés átprogramozásából származhatnak (1. ábra). Egy
adott gén szekvenciájában az allélvariánsok szekvenálással vagy
célzott genomhiányok indukálásával (Targeting Induced Local Lesions In
Genomes – TILLING) határozhatók meg. Így például a szárazságstresszben
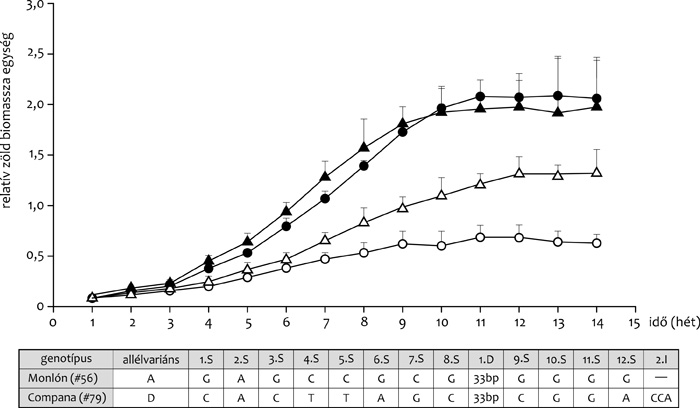
szerepet játszó egyik árpagén, a LEA-fehérjét kódoló HvA1 gén esetében
kapcsolat mutatható ki a haplotípus és a vízhiány okozta pixel alapú
zöldfelszín csökkenése között, mint ezt az 5. ábra szemlélteti.
A toleráns variánsban a D haplotípus található, míg a szenzitív
genotípusra az A haplotípus jellemző.
Napjainkban fokozott figyelem kíséri a genomikai
lenyomatfejlődés (genomic imprinting) biológiai szerepét. Az
endospermium szövet kialakulását szabályzó folyamatok kitűnő példát
szolgáltatnak erre a jelenségre. A magvak képződése feltételezi
bizonyos gének szülői, anyai vagy apai eredet szerinti eltérő
kifejeződését. Ezt a kiegyensúlyozatlan genetikai állapotot nevezik
genetikai lenyomatnak, ami nem a DNS-szekvenciában kódolt, hanem
epigenetikus folyamatok során jön létre, mint a hisztonfehérjék
módosítása vagy a DNS-metiláció. Az epigenetikus jelenségek
visszavezethetők a kromatin aktív, génkifejeződést megengedő vagy
zárt, a gének elhallgatását okozó állapotával. A Polycomb Group (PcG)
fehérjék mint epigenetikus tényezők a növényekben is megtalálhatók, és
részt vesznek a sejtek típusának meghatározásában, illetve az
osztódási vagy differenciálódási folyamatok közötti átmenet
szabályozásában. Az Arabidopsis mutánsok között találhatók az ún. FIS-
(Fertilization Independent Seed) gének hibái. Az ilyen PcG-gének
funkcióvesztése következtében endospermium-képződés indul meg annak
ellenére, hogy nem történik meg a megtermékenyítés.
4. Új génkombinációk születése izolált gének beépítésével: a
GM-növények térhódítása
és a hazai tiltás
Az izolált agronómiai gének funkcionális jellemzése legmegbízhatóbban
úgy végezhető el, ha a géneket beépítik a donor vagy idegen faj
genomjába, és megvizsgálják az okozott fenotípusos változásokat, az új
tulajdonságok öröklődését. Ezt a műveletsort nevezik genetikai
transzformációnak, az új genotípust cisz-, illetve transzgenikus
növénynek. A genetikai módosításból származó GM-növény megnevezés
szakmaiatlan, mert genetikai módosítás a keresztezés vagy a szelekció
is. Minthogy minden termesztett növényünk genetikailag módosul a
nemesítés során, indokoltabb géntechnológiával nemesített növényekről
beszélni. Magyarországon az utóbbi időkig több
kutatóhelyen is sikeres K+F-projekteket valósítottak meg, valamint
összefoglaló tanulmánykötetek készültek a növényi GMO-k jelentőségéről
az alapkutatásban és a növénytermesztési gyakorlatban: Géntechnológia
és gazdasági növényeink (Dudits, 2007); Zöld géntechnológia és
agrárinnováció (Dudits, 2009); Genetikailag módosított élőlények
(GMO-k) a tények tükrében: magyar fehér könyv (Balázs et al., 2011).
A 6. ábra tanúsága
szerint a növényi GMO-k megállíthatatlanul terjednek, és meghatározó
növénytermesztési technológiákat alapoznak meg. A többletjövedelem még
európai vonatkozásban is számottevő lehetne, ha a gazdálkodóknak
megengednék, hogy GM-kukoricát, -gyapotot, -szójababot, -olajrepcét és
-cukorrépát termesszenek ott, ahol ez agronómiailag indokolt,
nyereségük a becslések szerint évi 443–929 millió euróval emelkedne
(Park et al., 2011). A környezeti és egészségügyi hatásokat tekintve
egymásnak ellentmondó érvelések tömegével találkozhatunk. A média
aktív szerepvállalásának köszönhetően a tudományos érvek jelentősége
eltörpül a szenzációhajhász álhírek árnyékában. Idehaza igen hatásos
volt az emberek félretájékoztatása. Azokat a szakembereket, akik
tudományos munkásságuk tapasztalataival próbálják a tévedéseket
korrigálni, igaztalanul a multinacionális cégek bérenceiként bélyegzik
meg. Ezért a személyeskedő viták közepette talán hihetünk annak a
negyvenegy svéd tudósnak, akik állásfoglalásukkal kívánták felhívni a
figyelmet az európai helyzet visszásságaira (URL2 Hírlevél 2011/10).
Emlékeztetnek arra, hogy uniós forrásokból ötszáz független
kutatócsoport 300 millió eurót kapott a környezeti és egészségi
kockázatok vizsgálatára. A következtetés egyértelmű: a GM-technológia
önmagában nem veszélyesebb, mint a hagyományos nemesítés.
Figyelmen kívül hagyva a szakmai érveket és a
magyar gazdák hosszú távú érdekeit, a törvényhozás szélsőséges döntést
hozott, amikor az Alaptörvényben az alábbi géntechnológia-ellenes
betoldást szavazta meg:
Magyarország Alaptörvénye • Szabadság és felelősség
fejezet, 11. oldal, XX. cikk • Mindenkinek joga van a testi és lelki
egészséghez.
Az (1) bekezdés szerinti jog érvényesülését
Magyarország genetikailag módosított élőlényektől mentes
mezőgazdasággal, az egészséges élelmiszerekhez és az ivóvízhez való
hozzáférés biztosításával, a munkavédelem és az egészségügyi ellátás
megszervezésével, a sportolás és a rendszeres testedzés támogatásával,
valamint a környezet védelmének biztosításával segíti elő.”
A fenti megfogalmazás, függetlenül az élő szervezet
fajától, a hasznosítás módjától, és a transzgén által kódolt
tulajdonságtól, minden GM-szervezetet (GMO) a testi és lelki
egészséget veszélyeztető terméknek tekint. Az Alaptörvény szellemét
mélyen sérti ez a tudománytalan megállapítás. Bármely élelmiszerként
fogyasztott élő szervezet biztonsága annak tulajdonságaitól függ, és
nem attól, hogy génállományának kialakulása milyen genetikai
folyamatokkal történt. A törvényhozó szándék külön logikátlansága,
hogy csak a magyar földön, magyar gazda által előállított GMO-tól óvja
az emberek testi és lelki egészségét, míg megengedő azzal a
GM-termékkel szemben, amelyet importálunk. Miért nem hasznosíthatjuk
azokat a GMO-kat, amelyek gazdaságosabb bioenergia előállítását teszik
lehetővé? A saját érvényes törvényeinkkel és az uniós szabályokkal is
több ponton ütközik ez a végszavazás előtti pillanatokban tett
szövegmódosítás, mint azt Hetényi Kinga jogi szakvéleményében elemzi
(URL3, Hírlevél 2011/9).
Mostanáig az Európai Unióban két GM-növény
termesztését engedélyezték, míg 40 termék behozatalára és
forgalmazására van lehetőség. Ha a magyar növénytermesztés igényeit
tekintjük, az engedélyezés különböző szakaszaiban lévő GM-növények
közül a kukoricabogár-rezisztens hibridek, a fitoftóra (Phytophthora
infestans) nevű gombával szemben ellenálló burgonya használata
jelentene gazdasági és környezetvédelmi előnyt. Látva a nemzetközi
tudományos kutatás eredményeit, a folyamatban lévő fejlesztések
irányait és méreteit, nem kételkedhetünk abban, hogy a
géntechnológiára alapozott nemesítés kikerülhetetlen integráns része a
fajta-előállításnak. A magyar agrárium érdeke ki fogja kényszeríteni a
realitások tudomásulvételét, és az ennek megfelelő törvényi háttér
megteremtését.
Kulcsszavak: növénynemesítés, géntechnológia, képalkotás, genetikai
program, környezet, szárazságtűrés, haplotípus, GM-növényfajták,
Alaptörvény
IRODALOM
Balázs Ervin – Dudits D. – Sági L.
(szerk.) (2011): Genetikailag módosított élőlények (GMO-k) a tények
tükrében: magyar fehér könyv. Barabás Zoltán Biotechnológiai
Egyesület, Szeged •
WEBCÍM >
Cseri András – Sass L. – Törjék O. et al.
(2012): Phenomics Technologies Combined with Haplotyping in the
Analysis of Drought Responses of Barley Genotypes. (submitted)
Dudits Dénes (2004): A növények szexuális
életének molekuláris titkai. Mindentudás Egyeteme 4. kötet. 109–126. •
WEBCÍM >
Dudits Dénes (szerk.) (2007):
Géntechnológia és gazdasági növényeink. Magyar Tudomány. 4, 402–462. •
./07apr.html
Dudits Dénes (szerk.) (2009): Zöld géntechnológia és agrárinnováció.
Gazdafórum az Akadémián. Barabás Zoltán Biotechnológiai Egyesület,
Szeged. •
WEBCÍM >
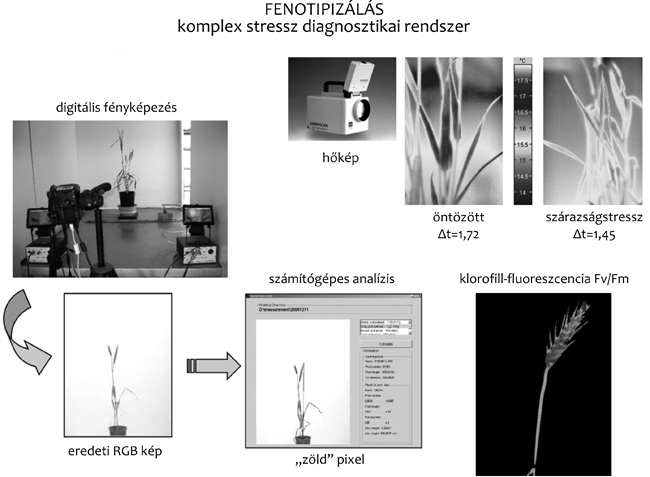
Hartmann, Anja – Czauderna, T. – Hoffmann,
R. et al. (2011): HTPheno: An Image Analysis Pipeline for
High-Throughput Plant Phenotyping. BMC Bioinformatics. 12, 148–156. •
WEBCÍM >
Houle, David – Govindaraju, D. R. –
Omholt, S. (2010) Phenomics: The Next Challenge. Nature Reviews
Genetics. 11, 855–866.
James, Clive (2011): Global Status of
Commercialized Biotech/GM Crops. 2010. ISAAA Brief 42. ISAAA, Ithaca,
NY
Park, Julian – McFarlane, I. – Phipps, R.
et al. (2011): The Impact of the EU Regulatory Constraint of
Transgenic Crops on Farm Income. New Biotechnology. 28, 4, 396–406.
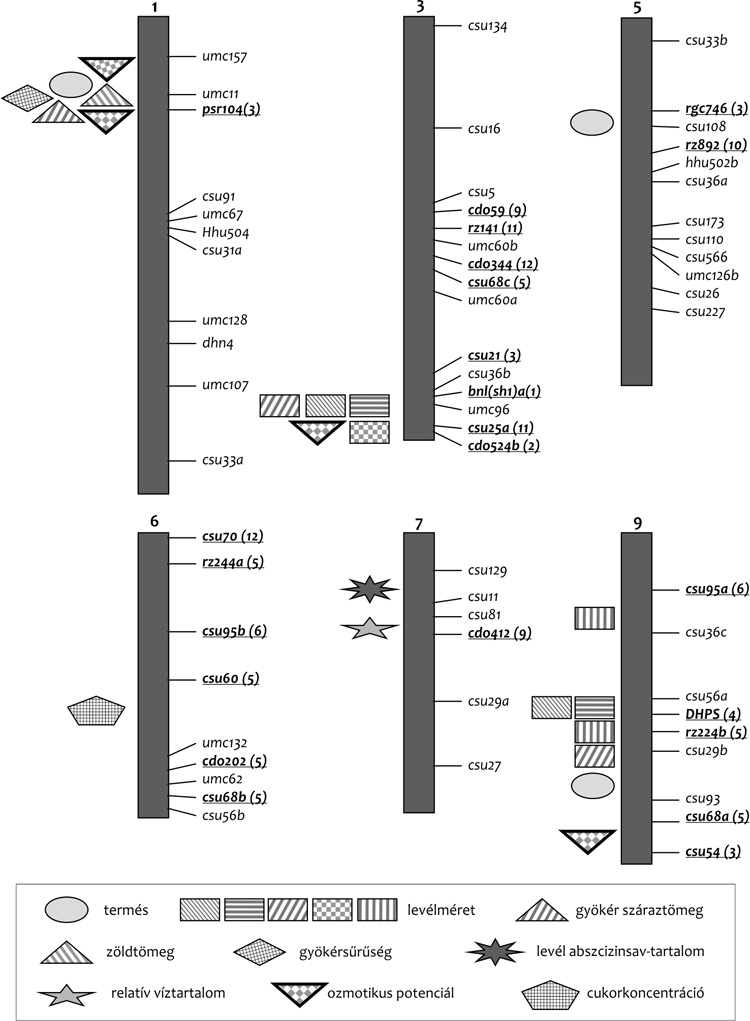
Rahman, H. – Pekic, S. – Lazic-Jancic, V.
et al. (2011): Molecular Mapping of Quantitative Trait Loci for
Drought Tolerance in Maize Plants. Genetics and Molecular Research.
10, 2, 889–901. •
WEBCÍM >
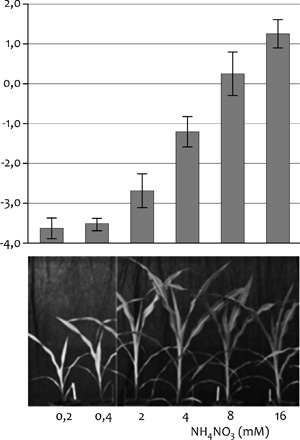
Yang, Xiaofeng S. – Wu, J. – Ziegler, T.
E. et al. (2011): Gene Expression Biomarkers Provide Sensitive
Indicators of in Planta Nitrogen Status in Maize. Plant Physiology.
157, 4, 1841–1852. •
WEBCÍM >
URL1
URL2
URL3
|