|
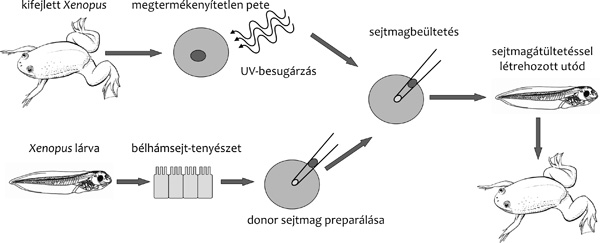
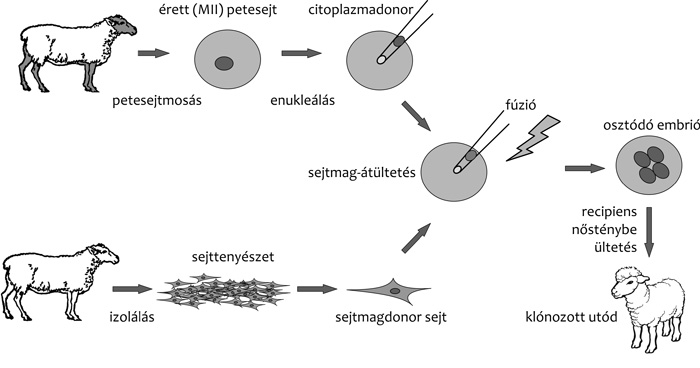
A sejtmagátültetéses klónozás alaptechnológiája nem
sokat változott a kezdetek óta. Minden faj esetén egy donorsejt
sejtmagjának preparálásával és egy megtermékenyítetlen petesejt –
esetleg zigóta, vagyis a még nem osztódott, korai egysejtes embrió –
enukleálásával, majd e két komponens valamilyen módszerrel történő
összeolvasztásával és/vagy aktiválásával folytatódik. Az osztódásnak
induló embriók azután – fajonként eltérő időpontban – recipiens
nőstényekbe kerülnek embrióbeültetéssel. A valódi különbségek az
egyes technikai lépések kivitelezésében keresendők (mint például
időzítés, oldatok összetétele, enukleálás vagy a fuzionáltatás
mikéntje stb.), amelyek egyrészt a módszer hatékonyságának javítása
érdekében történtek, másrészt az egyes fajok speciális embrionális
és/vagy hormonális karakteréhez igyekeznek mindinkább hasonlítani,
ezzel is biztosítva a nagyobb hatékonyságot. A sejtmagátültetéses
klónozás technológiai változásait több kiváló tanulmány is
feldolgozta, mind magyar (Dinnyés, 2004; Solti, 2004), mind pedig
angol nyelven (Wakayama, 2007; Dinnyés et al., 2008; Wilmut et al.,
2002, 2008); a terjedelmi korlátok miatt itt mi ennek részleteit nem
ismertetjük.
Ki kell térnünk azonban még egy nagyon fontos
momentumra, amely az őssejtkutatást érinti. Ez pedig nem más, mint a
sejtmagátültetéssel nyert embrionális őssejtek megjelenése. Az
embrionális őssejtekről e kötet későbbi tanulmányai részletesen
írnak majd, így itt csak a klónozott embriókból előállított
embrionális őssejtek jelentőségére térnénk ki. Az embrionális
őssejtek blasztociszta stádiumú embriók ún. belső sejtcsomójából
(Inner cell mass – ICM) izolált pluripotens sejtek (a fogalom
magyarázatát lásd később). Ezek az őssejtek optimális in vitro
körülmények között korlátlan ideig fenntarthatók, azonban mind in
vitro – Petri-csészében – mind pedig in vivo – recipiens embrióba
ültetve és kimérákat létrehozva – körülmények között képesek
differenciálódásra, a szervezet összes sejttípusának létrehozására.
Jelentőségük a célzott genetikai módosítás és terápiás célú
szövetregeneráltatás terén egyaránt nagyon jelentős. E sejtek
izolálása azonban számos faj esetén igen bonyolult és nagyon
alacsony hatékonysággal működő módszer. Megfigyelték azonban, hogy
sejtmagátültetéssel létrehozott embriók blasztocisztáit felhasználva
azokból nagyobb hatékonysággal lehet embrionális őssejteket
előállítani. Ezek az ún. klónozott embrionális őssejtek, mások
(Brambrink et al., 2006; Wakayama et al., 2006) és saját (Kobolák et
al., 2012a; Kobolák et al., 2012b) állatkísérletes eredményeink
szerint, nem térnek el tulajdonságaikban a hagyományos,
megtermékenyítéssel nyert blasztocisztákból izolált sejtvonalaktól.
A technológia humán vonatkozása azért nagyon jelentős, mert ez az
ún. terápiás célú klónozás nem más, mint egy differenciálódott testi
sejt pluripotens őssejtté történő alakítása. Vagyis egyedi,
személyre szabott őssejtek létrehozása is lehetséges anélkül, hogy
humán embriókat kellene felhasználni az embrionális őssejtek
létrehozására. Ezzel az eljárással testi sejtekből kiindulva,
„csupán” megtermékenyítetlen petesejtekre van szükség embriók
helyett az őssejtek előállításához. A továbbiakban látni fogjuk,
hogy az átprogramozás azonban már nem csak sejtmagátültetéssel
valósítható meg, így az oly sok etikai problémát is felvető
embrionális eredetű őssejtek és a klónozás technológiája is
megkerülhető.
Újraprogramozás sejtmagátültetés nélkül
Tudománytörténeti összefoglalónkban nem kisebb
feladatra vállalkoztunk, mint hogy bemutassuk a
sejtmagátültetéssel történő újraprogramozáson kívüli
sejtújraprogramozási módszerek fejlődésének mérföldköveit is.
Ezelőtt azonban még néhány fogalmat feltétlenül tisztáznunk kell,
habár e kötet további tanulmányai bőséges magyarázattal szolgálnak
majd az őssejtek és pluripotencia kérdéskörét illetően, a téma
legfontosabb fogalmait a 2. táblázatban
foglaltuk össze. Ahhoz azonban, hogy e terület eredményeit
értékelhessük, meg kell adnunk a pluripotencia definícióját.
Pluripotencia alatt a sejtek azon tulajdonságát értjük, hogy képesek
korlátlan számú osztódásra, miközben egy időben képesek önmaguk
reprodukálására és differenciálódott utódsejtek létrehozására is
(aszinkron sejtosztódás), és képesek – bizonyos körülmények között
és szignálok hatására – bármilyen sejt és szövet létrehozására, ide
értve mindhárom csíralemez (ekto-, mezo- és endoderma) sejtjeit és
az ivarsejteket is. Más szóval egy sejtet akkor nevezünk pluripotens
őssejtnek, ha az a teljes egyedfejlődési program „lefuttatására”
képes. Tehát nem csak DNS-tartalmában hordozza azt, hiszen azt az
előbbiekben a sejtmagátültetés kapcsán már tisztáztuk, hogy ez az
információ minden eukarióta sejt sajátja, hanem az információ olyan
„epigenetikai” státusban is van, hogy az kifejeződésre is jut.
Sejtkivonatok alkalmazása
Az egyik ilyen lehetőséget az egyes sejtek extraktumainak
alkalmazása jelenti, amikor valamely sejt – általában őssejt vagy
karcinómasejtek – valamilyen technológiával – általában
homogenizálás, centrifugálás és szűrés – készített kivonatait
használva az átalakítandó sejtekhez adjuk a kivonatot. A kivonatban
található molekulák – elsősorban fehérjék és más makromolekulák –
azután a recipiens sejtekben olyan változást idéznek elő, amely
aktivizálja az ún. pluripotens állapot fennmaradásáért felelős
genetikai programot, ezzel olyan, pluripotens vagy ahhoz közeli
állapotú utódsejtek jönnek létre a tenyészetben, amelyek azután az
amúgy őssejtekre jellemző differenciációs és fejlődési potenciállal
rendelkeznek (Taranger et al., 2005; Collas – Taranger 2006).
Philippe Collas és munkacsoportja 2005-ös és későbbi kísérleteinek
bírálói azt kifogásolják, hogy az átprogramozás sikere nagyban függ
a kiindulási sejtpopulációtól, amelyből az extraktum kinyerésre
kerül, így az újraprogramozás sikere és hatékonysága is kérdéses,
kísérletenként eltérő lehet. Az extraktum igen csekély mennyisége is
hatalmas kiindulási sejtpopulációt, méghozzá embrionális őssejteket
igényel, ami nagyon költségessé teszi az eljárást. A módszer sikere
sok kiindulási tényező változásától függ, és nem minden faj és
sejttípus esetén működik, továbbá a teljes genomra kiterjedő
újraprogramozás bizonyítékai nem meggyőzőek. Ezért ezen eljárások
alkalmazása nem nyert valódi teret az átprogramozásban.
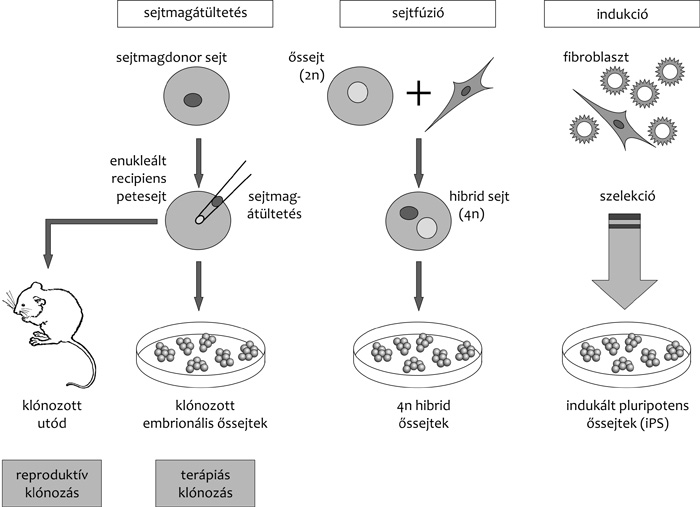
Sejtfúzió embrionális őssejtekkel
Pluripotens sejteket – embrionális karcinóma, ivari őssejtek vagy
embrionális őssejtek – differenciálódott testi sejtekkel
fuzionáltatva, a kiindulási őssejtekre „hasonlító” pluripotens
hibrid sejteket lehet létrehozni (fusion-mediated reprogramming –
FMR). Ezen utódsejtek azonban 4n kromoszómakészletűek,
vagyis tetraploidok. A tetraploid sejtekből specifikus módon
ugyan lehetséges nagyobb kromoszómarészletek eltávolítása, azonban
ez instabillá teheti a sejteket. A hatékony „diploidizálás” nem
megoldott. További probléma a sejtfúzió alacsony hatékonysága, így
ígéretes mivolta ellenére ezen eljárás sem hordoz terápiás
potenciált (Gan et al., 2007) (3. ábra).
Indukált átprogramozás
Az őssejtkutatás fejlődése, új őssejttípusok felfedezése, így
például a humán embrionális őssejtek izolálása 1998-ban (Thomson et
al., 1998), az ún. epiblaszt őssejtek (EpiSC) (Brons et al., 2007;
Tesar et al., 2007), vagy a multipotens felnőtt spermatogónium
őssejtek (multipotent adult spermatogonial stem cells – MASCs)
(Seandel et al., 2007) izolálása olyan összehasonlító vizsgálatokat
tett lehetővé, amelyek révén az őssejtek tulajdonságainak és a
pluripotencia mibenlétének egészen új aspektusai láthattak
napvilágot. A különböző pluripotens és multipotens sejtvonalak
karakterizálása, tenyésztési paramétereik vizsgálata és
összehasonlítása vezetett azután ahhoz a felismeréshez, hogy a humán
embrionális őssejtek (hESC), az epiblaszt őssejtek (EpiSC) és
spermatogónium őssejtek (MASCs) jobban hasonlítanak egymásra, mint
az egér embrionális őssejtekre (mESC). Mindez felvetette annak
lehetőségét, hogy a humán ESC-sejtek inkább epiblaszt eredetűek, nem
pedig ICM-eredetűek, mint az egérsejtek. Mindez azt is jelenti, hogy
e három őssejtfajta a pluripotencia más szintjén áll, mint az egér
ESC-sejtek. Ezek az eredmények egy egészen új gondolat
megszületéséhez vezettek, miszerint a pluripotencia megfelelő
faktorok aktiválása révén „előidézhető” lenne. Ennek vizsgálatára
2006-ban a most Nobel-díjat kapott Jamanaka és kollégája, Takahasi
Kazutosi (Kazutoshi Takahashi) újszerű kísérlettel álltak elő.
Rendszerükben négy gént, a pluripotencia fenntartásáért felelősnek
tartott ún. kulcsgéneket, név szerint Oct4, Sox2, valamint két
onkogént, a Klf4 és c-myc géneket juttatták be fibroblaszt –
embrionális és felnőtt – sejtekbe transzdukcióval. Ezt követően a
sikerrel transzdukált klónokban az Oct4 egyik targetgénjének, az
Fbx15 gén aktiválódását keresve pluripotens sejtpopulációt hoztak
létre, amelyet indukált pluripotens őssejteknek (induced pluripotens
stem) vagy iPS-sejteknek neveztek el (Takahashi – Yamanaka, 2006)
(3. ábra). Sejtjeik pluripotens stádiuma a virális promóterről
átíródó transzkriptumok jelenlétét követelte meg, az endogén Oct4 és
Nanog gének vagy nem is expresszáltak, vagy azok promotereinek
aktivitása nagyon alacsony maradt. Amikor az előzőekkel azonos módon
végzett transzdukció után az Oc4-et endogén expresszáló (Oct4-iPS)
vagy Nanogot expresszáló (Nanog-iPS) sejteket szelektáltak, akkor
azonban már valódi, az ESC-sejtekkel megegyező tulajdonságú
iPS-sejteket sikerült izolálni (Okita et al., 2007; Wernig et al.,
2007). Ezek a sejtek a pluripotencia minden kritériumát
teljesítették, egérben képesek voltak embrióba ültetve kimérákat
létrehozni, kolonizálva azok gonádját, így ivari kiméra utódokat
eredményezni. Sőt mi több, az ún. tetraploid kiméra eljárás során
ezen iPS-sejtek egy lépésben képesek voltak teljes iPS-sejt eredetű
egyed létrehozására is, ahol a tetraploid embrió csupán a placenta
kialakulásában vett részt. Mindez azt jelenti, hogy e külső faktorok
ektopikus expressziója képes volt indukálni olyan epigenetikai
változásokat – kromatin struktúra és DNS-metilációs változások – a
sejtekben, amely azok átprogramozásához vezetett. Ezzel megszületett
egy egészen új, immár embrió- és sejtmagátültetés-mentes módszer a
differenciálódott sejtek újraprogramozására.
A humán iPS-sejtek létrehozása sem váratott sokat
magára, ebben Jamanaka kutatócsoportja ismét az élen járt (Takahashi
et al., 2007). A továbbiakban viharos gyorsasággal szaporodott a
közlemények száma, ahol iPS-sejteket izoláltak különféle
sejtforrásokból. Több különböző faktor is kipróbálásra került az
eredeti négy helyett, és megoldottá vált az onkogén-, vírus- és
antibiotikum-rezisztencia-mentes átprogramozás is. Így elérhetővé
vált a genetikai módosításmentes iPS-előállítás, ami a technológia
génterápiás alkalmazásának egyik előfeltétele (Yamanaka, 2012). A
kötet további tanulmányaiban bővebb értékelést olvashatnak a
különböző őssejtekről, többek között az iPS-sejtekről és e
tudományterület új áttöréseiről, így tudománytörténeti
áttekintésünkben az eredmények részletes ismertetésébe nem
bocsátkozunk.
Összefoglalás
Az újraprogramozás biológiai és orvostudományi jelentősége hatalmas.
A személyre szabott gyógyítás terén olyan új távlatok nyíltak meg
ezen eljárások alkalmazása révén, amely számos, korábban
gyógyíthatatlannak tartott betegség esetén áttörést jelent. Bátran
kijelenthetjük, hogy soha olyan gyors fejlődésen és átalakuláson nem
ment még át e tudományterület, mint amelyet az újraprogramozás
felfedezése hozott. Meggyőződésünk, hogy e terület fejlődése,
különösen az iPS-technológia területén, további forradalmi
változásokat hoz majd a gyógyszerkutatás és gyógyítás területén is.
Kulcsszavak: sejtmagátültetés, újraprogramozás, epigenetika,
pluripotencia, embrionális őssejt, indukált pluripotens őssejt
IRODALOM
Beyhan, Z. – Iager, A. E. – Cibelli, J. B.
(2007): Interspecies Nuclear Transfer: Implications for Embryonic
Stem Cell Biology. Cell Stem Cell. 1, 5, 502–512.
Brambrink, T. – Hochedlinger, K. – Bell,
G. – Jaenisch, R. (2006): Es Cells Derived from Cloned and
Fertilized Blastocysts Are Transcriptionally and Functionally
Indistinguishable. Proceedings of the National Academy of Sciences
of the USA. 103, 4, 933–938.
Brons, I. G. – Smithers, L. E. – Trotter,
M. W. – Rugg-Gunn, P. – Sun, B. – Chuva De Sousa Lopes, S. M. –
Howlett, S. K. – Clarkson, A. – Ahrlund-Richter, L. – Pedersen, R.
A. et al. (2007): Derivation of Pluripotent Epiblast Stem Cells from
Mammalian Embryos. Nature. 448, 7150, 191–195.
Collas, P. – Taranger, C. K. (2006):
Epigenetic Reprogramming of Nuclei Using Cell Extracts. Stem Cell
Reviews. 2, 4, 309–317.
Dinnyés A. (2004): Őssejtek és a klónozás
lehetőségei. Magyar Tudomány. 3, 292–297.
Dinnyés A. – Tian, X. C. – Yang, X.
(2008): Epigenetic Regulation of Foetal Development in Nuclear
Transfer Animal Models. Reproduction in Domestic Animals. 43, Suppl.
2: 302–309.
Gan, Q. – Yoshida, T. – McDonald, O. G. –
Owens, G. K. (2007): Concise Review: Epigenetic Mechanisms
Contribute to Pluripotency and Cell Lineage Determination of
Embryonic Stem Cells. Stem Cells. 25, 1, 2–9.
Gilbert, S. F. (2006): Developmental
Biology. Sinauer Associates Inc. Publishers, Sunderland, Mass.
Gurdon, J. B. (2006): From Nuclear
Transfer to Nuclear Re-programming: The Reversal of Cell
Differentiation. Annual Review of Cell and Developmental Biology.
22, 1–22.
Hwang, I. – Jeong, Y. W. – Kim, J. J. –
Lee, H. J. – Kang, M. – Park, K. B. – Park, J. H. – Kim, Y. W. –
Kim, W. T. –
Shin, T. et al. (2012): Successful Cloning
of Coyotes through Interspecies Somatic Cell Nuclear Transfer Using
Domestic Dog Oocytes. Reproduction Fertility and Development.
Kobolák J. – Horsch, M. – Geissler, S. –
Mamo, S. – Beckers, J. – Dinnyés A. (2012a): Comparative Analysis of
Nuclear Transfer Embryo-derived Mouse Embryonic Stem Cells. Part II:
Gene Regulation. Cell Reprogramming. 14, 1, 68–78.
Kobolák J. – Mamo, S. – Rungsiwiwut, R. –
Ujhelly O. – Csonka E. – Hadlaczky G. – Dinnyés A. (2012b):
Comparative Analysis of Nuclear Transfer Embryo-derived Mouse
Embryonic Stem Cells. Part I: Cellular Characterization. Cell
Reprogramming. 14, 1, 56–67.
Okita, K. – Ichisaka, T. – Yamanaka, S.
(2007): Generation of Germline-Competent Induced Pluripotent Stem
Cells. Nature. 448, 7151, 313–317.
Seandel, M. – James, D. – Shmelkov, S. V.
– Falciatori, I. – Kim, J. – Chavala, S. – Scherr, D. S. – Zhang, F.
– Torres, R. – Gale, N. W. et al. (2007): Generation of Functional
Multipotent Adult Stem Cells from Gpr125+ Germline Progenitors.
Nature. 449, 7160, 346–350.
Solti L. (2004): Klónozás és génmódosítás:
szép új világ? Magyar Tudomány. 2, 1980–207.
Takahashi, K. – Tanabe, K. – Ohnuki, M. –
Narita, M. – Ichisaka, T. – Tomoda, K. – Yamanaka, S. (2007):
Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by
Defined Factors. Cell. 131, 5, 861–872.
Takahashi, K. – Yamanaka, S. (2006):
Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult
Fibroblast Cultures by Defined Factors. Cell. 126, 4, 663–676.
Taranger, C. K. – Noer, A. – Sorensen, A.
L. – Hakelien, A. M. – Boquest, A. C. – Collas, P. (2005): Induction
of Dedifferentiation, Genomewide Transcriptional Programming, and
Epigenetic Reprogramming by Extracts of Carcinoma and Embryonic Stem
Cells. Molecular Biology of the Cell. 16, 12, 5719–5735.
Tesar, P. J. – Chenoweth, J. G. – Brook,
F. A. – Davies, T. J. – Evans, E. P. – Mack, D. L. – Gardner, R. L.
– McKay, R. D. (2007): New Cell Lines from Mouse Epiblast Share
Defining Features with Human Embryonic Stem Cells. Nature. 448,
7150, 196–199.
Thomson, J. A. – Itskovitz-Eldor, J. –
Shapiro, S. S. – Waknitz, M. A. – Swiergiel, J. J. – Marshall, V. S.
– Jones, J. M. (1998): Embryonic Stem Cell Lines Derived from Human
Blastocysts. Science. 282, 5391, 1145–1147.
Wakayama, S. – Jakt, M. L. – Suzuki, M. –
Araki, R. – Hikichi, T. – Kishigami, S. – Ohta, H. – Van Thuan, N. –
Mizutani, E. – Sakaide, Y. et al. (2006): Equivalency of Nuclear
Transfer-derived Embryonic Stem Cells to Those Derived from
Fertilized Mouse Blastocysts. Stem Cells. 24, 9, 2023–2033.
Wakayama, T. (2007): Production of Cloned
Mice and Es Cells from Adult Somatic Cells by Nuclear Transfer: How
to Improve Cloning Efficiency? Journal of Reproduction and
Development. 53, 1, 13–26.
Wernig, M. – Meissner, A. – Foreman, R. –
Brambrink, T. – Ku, M. – Hochedlinger, K. – Bernstein, B. E. –
Jaenisch, R. (2007): In Vitro Reprogramming of Fibroblasts into a
Pluripotent Es-Cell-Like State. Nature. 448, 7151, 318–324.
Wilmut, I. – Schnieke, A. E. – McWhir, J.
– Kind, A. J. – Campbell, K. H. S. (1997): Viable Offspring Derived
from Fetal and Adult Mammalian Cells. Nature. 385, 6619, 810–813.
Wilmut, I. – Beaujean, N. – De Sousa, P.
A. – Dinnyés, A. – King, T. J. – Paterson, L. A. – Wells, D. N. –
Young, L. E. (2002): Somatic Cell Nuclear Transfer. Nature. 419,
6907, 583–586.
Wilmut, I. – Sullivan, G. – Taylor, J.
(2008): A Decade of Progress since the Birth of Dolly. Reproduction,
Fertility and Development. 21, 1, 95–100.
Yamanaka, S. (2012): Induced Pluripotent
Stem Cells: Past, Present, and Future. Cell Stem Cell. 10, 6,
678–684.
Zhu, Z. Y. – Sun, Y. H. (2000): Embryonic and Genetic Manipulation
in Fish. Cell Research. 10, 1, 17–27.
LÁBJEGYZETEK
1 A különböző források
különböző évszámokat adnak meg, az itt közölt dátumok Scott F.
Gilbert Developmental Biology című munkája alapján kerülnek közlésre
(Gilbert, 2006).
<
2 A születési évszám nem
feltétlenül egyezik meg a publikálás évével, illetve a publikáció
megjelenésének évével. Ebből adódóan az egyes források között
eltérések lehetnek. Ezért összefoglaló táblázatunkban az egyes
eredmények tudományos közleményeinek referenciáját is megadtuk, a
közzététel évével. Több esetben azonban, tudományos közlemény
hiányában, csak a médiában történő bejelentésre hagyatkozhattunk.
<
|