|
|
Bevezetés
A természetvédelmi munka jelentős hányadát teszik ki a
veszélyeztetett fajok védelmével kapcsolatos feladatok.
Megtervezésükben egyre nagyobb szerepet játszik a kérdéses fajok
genetikai variabilitásának ismerete, különös tekintettel annak
szerkezetére, a genetikai differenciálódás mintázatára. A genetikai
differenciálódás szintjét tekintve a természetvédelmi genetikai
szakirodalom két kulcsfogalmat használ: evolúciósan szignifikáns
egység (Evolutionary Significant Unit – ESU) és konzervációs egység
(Conservation Unit – CU). Bár a fogalmak használata nem teljesen
egyöntetű a különböző szakcikkekben, az ESU kritériumai
többé-kevésbé jól definiáltak (Moritz, 1994; Crandall et al., 2000).
Azokat a populációkat/ populációcsoportokat tekinthetjük önálló
evolúciós egységeknek, amelyek egyértelmű genetikai elkülönülést
mutatnak a közel rokon populációcsoportoktól, esetleg részleges
reproduktív izoláció is megjelenik közöttük. Az ESU a genetikai
differenciálódás mellett ökológiai elkülönülést is mutat a többi
populációtól, ami földrajzi izolációval is párosulhat. Továbbá az
evolúciósan szignifikáns egységek kölcsönösen monofiletikusak.
Ezeknek a kritériumoknak leginkább önálló taxonok, elsősorban fajok,
alfajok, esetleg ökológiai vagy földrajzi rasszok felelnek meg. A CU
azonban a gyakorlati természetvédelmi munkában jobban használható
fogalom, amennyiben azokat a populációkat /populációcsoportokat
jelöli, amelyek jelentős mértékben hozzájárulnak a kérdéses faj
genetikai diverzitásához, így speciális figyelmet, élőhelykezelést
érdemelnek. A konzervációs egységet alkotó populációknak is fontos
ismérvük tehát a többi populációtól való genetikai differenciálódás,
valamint a legalább részleges ökológiai elkülönülés. A
természetvédelmi genetikai vizsgálatoknak egyik fontos célkitűzése,
hogy a genetikai differenciálódás mintázata alapján megállapítsák a
védett és veszélyeztetett fajok populációcsoportjainak státuszát az
ESU- és CU-kategóriák tekintetében. A különböző fajok nagyon eltérő
differenciálódási mintázatot mutatnak, tehát változó az is, hogy
genetikai struktúrájukat az evolúciósan szignifikáns vagy a
konzervációs egységek megléte jellemzi. A fenti fogalmakat a
továbbiakban négy esettanulmányon keresztül mutatjuk be.
Sötét hangyaboglárka (Maculinea nausithous)
A Kárpát-medence környéki sötét hangyaboglárka populációk
tanulmányozása során a minták három földrajzi régió (Dunántúl,
Erdély, Bukovina) tizenkét populációjából származtak, összesen 463
egyedben történt az enzim polimorfizmus (= az egyes enzimeket kódoló
genetikai egységek sokféleségének) vizsgálata. Tizenhét
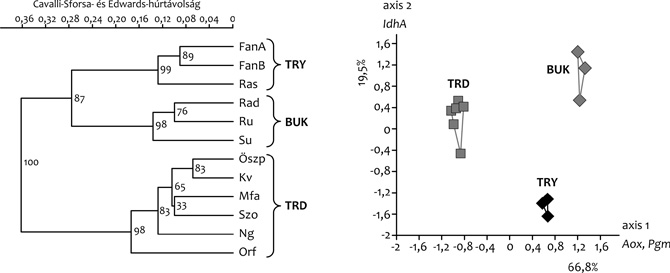
enzim-lókuszon elemeztük a populációk allélfrekvencia-adatait, majd
ezek segítségével Cavalli-Sforza- és Edwards-húrtávolságokat
számoltunk, és UPGMA-dendrogramot szerkesztettünk. Eredményeink azt
mutatták, hogy a keleti (erdélyi és bukovinai), valamint a nyugati
(dunántúli) populációk között jelentős mértékű a differenciálódás
(1a ábra). Ezt támasztotta alá
a főkomponens-analízis eredménye is, ahol az első tengely mentén,
ami a teljes genetikai variancia 66,8%-át magyarázta, egyértelműen
elkülönült a két populációcsoportot jelképező pontfelhő (1b
ábra). A Bayes-féle klaszteranalízis (Pritchard et al., 2000)
eredménye azt mutatta, hogy két genetikai klaszter megléte a
legvalószínűbb a vizsgált mintákban: az 1. klaszter a dunántúli
populációkban volt jellemző, míg a 2. klaszter a keletiekben (Erdély
és Bukovina). Az erdélyi és a bukovinai populációk genetikai
különbsége akkor tárult fel, amikor azt tételeztük fel, hogy három
genetikai klaszter van jelen a populációkban. Ebben az analízisben a
három régiót egy-egy specifikus klaszter túlsúlya jellemezte.
Érdekes módon, a dunántúli egyedekben az erdélyi genetikai állomány
jelent meg szórványosan, míg a bukovinai alig. Ugyanakkor, mind az
erdélyi, mind pedig a bukovinai régió egyedei kölcsönösen nagyobb
mértékű keveredést mutattak. Eredményeink megfeleltek a faj
elterjedési területe alapján kialakított hipotézisünknek. A sötét
hangyaboglárka elterjedési területe ugyanis regionálisan
megszakított, amennyiben a Kárpát-medence középső területein a faj
nem fordul elő. A dunántúli populációk lényegében az elterjedési
terület nyugati részének peremén, míg a bukovinai populációk a
keleti területek peremén találhatók. Az erdélyi populációk
feltehetően az área keleti felének leszakadó maradványai. Úgy tűnik
tehát, hogy a sötét hangyaboglárka keleti és nyugati populációi
alfaji szinten különülnek el egymástól (Rákosy et al., 2010), vagyis
különböző evolúciós egységeknek tekinthetők.
Lápi tarkalepke (Euphydryas aurinia)
A Kárpát-medence környéki lápi tarkalepke populációk mintái négy
földrajzi régió (Szlovénia, Őrség, Közép-Dunántúl és Erdély) kilenc
populációjából származtak. Összesen 281 egyedben tanulmányoztuk a
variabilitás mértékét 18 enzimlókuszon. Az allélfrekvencia-adatokat
különböző statisztikai analíziseknek vetettük alá. Az eredmények
azonban csak részben voltak következetesen értelmezhetők. A
Cavalli-Sforza- és Edwards-húrtávolságok alapján szerkesztett
UPGMA-dendrogram azt mutatta, hogy az erdélyi (keleti) populáció
egyértelműen differenciálódott a többitől (nyugati populációk). Ezt
támasztotta alá a főkomponens-analízis eredménye is, ahol az erdélyi
populáció az első tengely mentén különült el a nyugatiaktól. Ez a
tengely a teljes genetikai variancia 53,7%-át magyarázta. Ugyanakkor
ezekben az analízisekben nem mutatkozott egyértelmű mintázat a
nyugati populációk genetikai struktúrájában. A Bayes-féle
klaszteranalízisben a klaszterek legvalószínűbb száma háromnak
adódott. Amikor a klaszterek eloszlását az egyedekben megvizsgáltuk,
akkor kiderült, hogy az erdélyi populációra a 3. klaszter a jellemző
(97%), ezekben az egyedekben a másik két klaszter alig fordult elő.
Ez az eredmény arra enged következtetni, hogy a nyugati populációk
egyedei gyakorlatilag nem keveredtek az erdélyiekkel. A dunántúli és
a szlovéniai populációkban viszont az 1. és 2. klaszter változó
gyakorisággal jelent meg. Az őrségi és a szlovéniai populációkra
elsősorban az 1. (66,2%), míg a közép-dunántúliakra a 2. klaszter
(78,4%) volt jellemző. A lápi tarkalepke európai elterjedése
annyiban hasonlít a sötét hangyaboglárkáéhoz, hogy ennek a fajnak az
elterjedésében is jelentős hiátus tapasztalható a Kárpát-medence
belső részén. A szlovéniai, az őrségi és a közép-dunántúli
populációk az área nyugati részének peremén helyezkednek el, míg az
erdélyi populáció a keleti área leszakadt szigetének tekinthető, bár
összeköttetése még nem világos. Ehhez a kelet-balkáni populációk
vizsgálata volna szükséges.
Az enzim polimorfizmus vizsgálatának eredményei
megfeleltek a faj elterjedési területe alapján várt
differenciálódási mintázatnak, megmutatták az erdélyi és a nyugati
populációk elkülönülését. Ám annak eldöntéséhez, hogy a
differenciálódás mértéke alapján a keleti és a nyugati populációk
önálló evolúciós egységeknek (alfajoknak) tekinthetők-e, több minta
volna szükséges elsősorban a faj áreájának keleti-délkeleti
részéből, például a Balkán-félszigetről. Érdekes viszont, hogy a
genetikai klaszterek alapján az őrségi és szlovéniai, valamint a
közép-dunántúli populációk között jelentős differenciálódást
tapasztaltunk. További vizsgálatokra, esetleg további markerekkel
történő analízisekre lenne szükség annak kimutatására, hogy a két
földrajzilag meglehetősen közeli régió populációi tekinthetők-e
különböző konzervációs egységeknek.
Kis apollólepke (Parnassius mnemosyne)
A kis apollólepke mintákat a Kárpát-medence öt régiójának
(Dunántúli-középhegység, Északi-középhegység, Körös-vidék,
Beregi-sík és Erdélyi-szigethegység) tizenkilenc populációjából
gyűjtöttük. Az 501 egyedben tizennégy enzimlókuszon határoztuk meg
az allélok eloszlását. Az allélfrekvencia-adatok alapján
Cavalli-Sforza- és Edwards-húrtávolságokat számítottunk, majd a
távolságmátrix segítségével UPGMA-dendrogramot szerkesztettünk.
Meglepő módon, a dendrogram csak a Körös-vidék két populációjának
elkülönülést mutatta. Ugyanakkor a főkomponens-analízisben az első
két tengely mentén négy régió differenciálódott
|
|
|
egymástól: a Dunántúli-középhegység,
Északi-középhegység, Körös-vidék és Beregi-sík + Erdély. Ez a két
tengely a genetikai variancia 42,9%-át magyarázta.
A kis apollólepke-populációk genetikai struktúrája
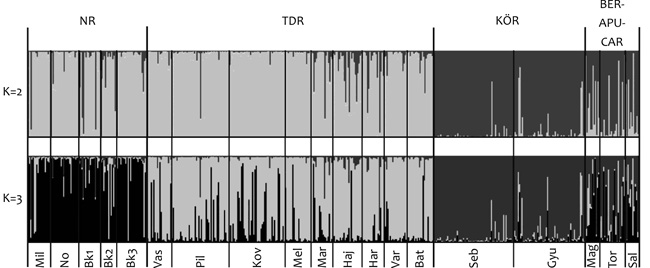
azonban a Bayes-féle klaszteranalízisben vált egyértelművé. A
genetikai klaszterek legvalószínűbb száma kettő volt. A két klaszter
egyedekben való megoszlása alapján három régió rajzolódott ki
(2. ábra): az Északi- és a
Dunántúli-középhegység egyedeiben az 1. klaszter volt szinte
kizárólagosan jelen, a Körös-vidéket a 2. klaszter döntő túlsúlya
jellemezte, míg a Beregi-sík és Erdély-régióban a két klaszter
70–30%-os megoszlást mutatott. Megvizsgáltuk azt a lehetőséget is,
hogy a populációkat három genetikai klaszter alkotja, aminek a
valószínűsége még szintén magas volt. A három klaszter feltételezése
a differenciálódás mintázatát annyiban tette világosabbá, hogy (1)
az Északi- és a Dunántúli-középhegység populációit különböző
klaszterek jellemezték; (2) a Beregi-sík és Erdély-régióban az
Északi-középhegységet jellemző 1. klaszter, és a Körös-vidéket
alkotó 3. klaszter keveredését figyeltük meg. Eredményeink jó
egyezést mutattak Paolo Gratton és munkatársai (2008) tanulmányával,
akik a mtDNS-szekvenciaadatok alapján azt feltételezték, hogy
Közép-Európa keleti részét a P. mnemosyne két fő irányból, az Alpok
DK-i pereme, illetve a Keleti-Kárpátok felől népesítette be.
Munkájukból kitűnik, hogy a Kárpát-medence benépesítésében
egyértelműen az utóbbi irányból történő volt a meghatározó. Ez a
genetikai ág is kettős tagolódást mutat. Keleti alága a
Keleti-Kárpátok felől nyomult be Erdélybe és a Kárpát-medence keleti
területeire, míg a nyugati alág a Déli-Kárpátok megkerülésével
jutott a Kárpát-medence középső és nyugati területeire, majd ez az
alág népesítette be Észak-Európa jelentős részét is. Az általunk
talált két genetikai klaszter valószínűleg azonos a Gratton és
munkatársai által leírt kelet-európai ág nyugati és keleti
alágaival. Ezt a feltételezést alátámasztja az is, hogy a
középhegységi populációk (1. klaszter) esetében a Mantel-teszt
nagyon szoros korrelációt (R=0,524; P<0,001) mutatott a genetikai és
a földrajzi távolságok között, ami egyértelmű bizonyítéka annak,
hogy ezeknek a populációknak a genetikai struktúráját a távolsággal
arányos izoláció jellemzi. Bár eszerint a Kárpát-medencei P.
mnemosyne populációk egy evolúciós egységet alkotnak, a Bayes-féle
klaszteranalízis eredményei alapján elkülönülő három régiót
mindenképpen önálló konzervációs egységnek kell tekintenünk.
A szürkés hangyaboglárka
nedvesréti ökotípusa (Maculinea alcon)
A hangyaboglárkák sajátságos életmenettel jellemezhető nappali
lepkék. A nőstények speciális tápnövényekre rakják petéiket. A
kikelő hernyók eleinte a tápnövény fejlődő magjaival táplálkoznak,
majd az éretlen termésekből kipotyogva várják, hogy különböző
Myrmica-fajok felfedezzék és adoptálják őket. Sikeres adoptálás
esetén a hernyók a specifikus hangyagazdák fészkeibe kerülnek, és
ott fejlődnek tovább, valamint ott is bábozódnak be. A
vizsgálatokban használt szürkés hangyaboglárka minták hat földrajzi
régió (Kiskunság, Mátra, Zempléni-hg., Szatmári-sík, Beregi-sík és
Erdély) kilenc populációjából származtak. Az enzim polimorfizmust
386 egyedben, tizenhat lokuszon tanulmányoztuk. A Bayes-féle
klaszteranalízisben a klaszterek legvalószínűbb száma háromnak
adódott. A klaszterek megoszlása alapján három „genetikai régió”
mutatkozott: Zempléni-hg. (3. klaszter: ~70%), Fülesd (Szatmári-sík;
2. klaszter: ~70%) és a viszonylag heterogén Kiskunság – Mátra –
Beregi-sík – Erdély csoport (1. klaszter: ~50%). Ha megvizsgáljuk,
hogy mi kapcsolhatja össze az egyes genetikai régiók populációit,
akkor azt tapasztaljuk, hogy a fő hangyagazda a különböző „genetikai
régiók” között eltér: a zempléni M. alcon populációk elsődlegesen a
My. vandelit parazitálják, a fülesdi populáció fő hangyagazdája a
My. slovaca, míg a harmadik régióban a szürkés hangyaboglárka
túlnyomórészt a My. scabrinodis fészkekben fejlődik.
A M. alcon hernyók a hangyagazda fészkében kakukkfióka módjára
élnek, vagyis hangyalárvaként viselkednek. Így a dolgozók etetik és
gondozzák őket. Ehhez alapvetően az szükséges, hogy a hernyók
mimikrije (hangyalárvák szagának, hangjának utánzása) nagyon pontos
legyen, vagyis kritikus a lokális adaptáció. Feltételezhetjük tehát,
hogy a különböző hangyagazdát hasznosító populációk adaptációja
eltérő, azaz közöttük divergens szelekció hatott. Ennek eredménye
úgy nyilvánul meg, hogy az ilyen hatásnak kitett lókuszon a
differenciálódás szintje magasabb, mint azokon, amelyeken csak
véletlen hatások (genetikai sodródás) érvényesülnek. Amikor a 16
vizsgált enzimlókusz között olyanokat kerestünk, amelyek kiugró
differenciálódást mutatnak, azt tapasztaltuk, hogy négy lókusz
esetében (Acon, Est, Hk és Mdh) a véletlen hatások alapján
várhatónál szignifikánsan (P<0,05) magasabb volt a differenciálódást
jellemző érték (fixációs index: FST). Különösen figyelemre méltó,
hogy a Bayes-féle analízisben kimutatott három klaszterben ezen a
négy lókuszon találtunk klaszterspecifikus allélokat. Eredményeink
alapján megállapítottuk, hogy a vizsgált M. alcon populációk egy
evolúciós egységet alkottak, ugyanakkor a különböző hangyagazdát
használó populációk, melyek között genetikai és ökológiai
differenciálódást is tapasztaltunk, önálló konzervációs egységeknek
tekinthetők.
Kitekintés
Az egyes fajok genetikai struktúráját számos tényező kölcsönhatása
alakítja ki. A fajok életmenete, szaporodási struktúrája és
demográfiai jellemzői mind meghatározzák a szelekciós folyamatok és
a véletlen hatások genetikai következményeit. Az elterjedési terület
földrajzi jellemzői, a populációk elterjedési akadályainak helye és
hatékonysága pedig a génáramlás intenzitását befolyásolják. Ezeken a
folyamatokon túlmenően a fajok történetisége, jégkorszaki
refúgiumaik száma és helye, valamint a jelenlegi áreájuk
kialakulásához vezető benépesedési utak mind hatással voltak és
vannak a genetikai differenciálódás (ESU) és a benépesített
élőhelyekhez való alkalmazkodás (CU) mintázatára. A fenti
esettanulmányok is világosan mutatják, hogy a fajok genetikai
struktúrája nagyon is különböző lehet. Ezért a gyakorlati
természetvédelmi munkában különösen fontos, hogy a veszélyeztetett
fajok védelmére kidolgozott programokat populációgenetikai
vizsgálatok eredményei alapján is megalapozottá tegyék.
Az eredmények nem születhettek volna meg Mester Valéria
laboratóriumi munkákban nyújtott segítsége nélkül. A minták
begyűjtésében több kolléga is részt vett: Ilonczai Zoltán, Kozma
Péter, Peregovits László, dr. Szabó Sándor és dr. V. Sipos Julianna.
A kutatást a MacMan EU FP6 projekt, az NKFP-3 B/ 023/2004, az OTKA
T030528, és K84071 pályázatok támogatták.
Kulcsszavak: evolúciósan szignifikáns egység, konzervációs
egység, zanótboglárka, lápi tarkalepke, lápi hangyaboglárka, kis
apollólepke
IRODALOM
Crandall, Keith A. – Bininda-Emonds, O. R.
P – Mace, G. M. – Wayne, R. K. (2000): Considering evolutionary
processes in conservation biology. Trends in Ecology & Evolution.
15, 290–295. doi:10.1016/S0169-5347(00)01876-0 •
WEBCÍM
Gratton, Paolo – Konopinski, M. K. –
Sbordoni, V. (2008): Pleistocene Evolutionary History of the Clouded
Apollo (Parnassius mnemosyne): Genetic Signatures of Climate Cycles
and a ‘Time-dependent’ Mitochondrial Substitution Rate. Molecular
Ecology. 17, 4248–4262. DOI: 10.1111/j.1365-294X.2008.03901.x
Moritz, Craig (1994): Defining
Evolutionarily-significant-Units for Conservation. Trends in Ecology
& Evolution. 9, 373–375. •
WEBCÍM
Pritchard, Jonathan K. – Stephens, M. –
Donnelly, P. (2000): Inference of Population Structure Using
Multilocus Genotype Data. Genetics. 155, 945–959. •
WEBCÍM
Rákosy László – Tartally, A. – Goia, M. –
Mihali, C. – Varga, Z. (2010): The Dusky Large Blue – Maculinea
nausithous (Bergsträsser, [1779]) in the Transylvanian Basin: New
Data on Taxonomy and Ecology. Nota Lepidopterologica. 33, 31-37. •
WEBCÍM
|
|