|
|
A Magyar Tudományban megjelent előző cikkem
(Podani, 2010) részletesen beszámolt az evolúció és a rendszertan
kapcsolatáról, és arról, hogy a biológia és a számítástechnika
fejlődésének köszönhetően ezeken a tudományterületeken milyen
lényeges változásokra kell odafigyelnünk. Mint a cím is jelezte, a
téma megkerülhetetlen a törzsfák és hasonló jellegű ábrázolások,
mint például a kladogram vagy az evolúciós fa tárgyalása nélkül,
hiszen ma már ezek szolgálnak az osztályozás alapjául. Néhány
példával illusztráltam, hogy a törzsfák és kladogramok értelmezése
számos buktatót rejt: a vizsgálatba bevont fajok és más taxonok
kapcsolatrendszerét igen könnyű félreérteni. A fák sokféleségének
témakörét természetesen nem merítettem ki, az alapfogalmak és a
köznyelv kapcsolatáról alig szóltam, számos történeti és elméleti
vonatkozást nem említettem – már csak a korlátozott oldalszám miatt
sem. Ebben a cikkben szeretnék néhány kiegészítést tenni – az
időközben született elméleti eredmények és több, nemrég megjelent
összefoglaló munka (például: Pietsch, 2012; Archibald, 2014)
figyelembevételével. A fő tanulságot a cikk címe is mutatja: nem
minden fa, ami annak látszik, s ami megfordítva is igaz lehet: sok
ábra a látszat ellenére is megfelel a fa egyes kritériumainak.
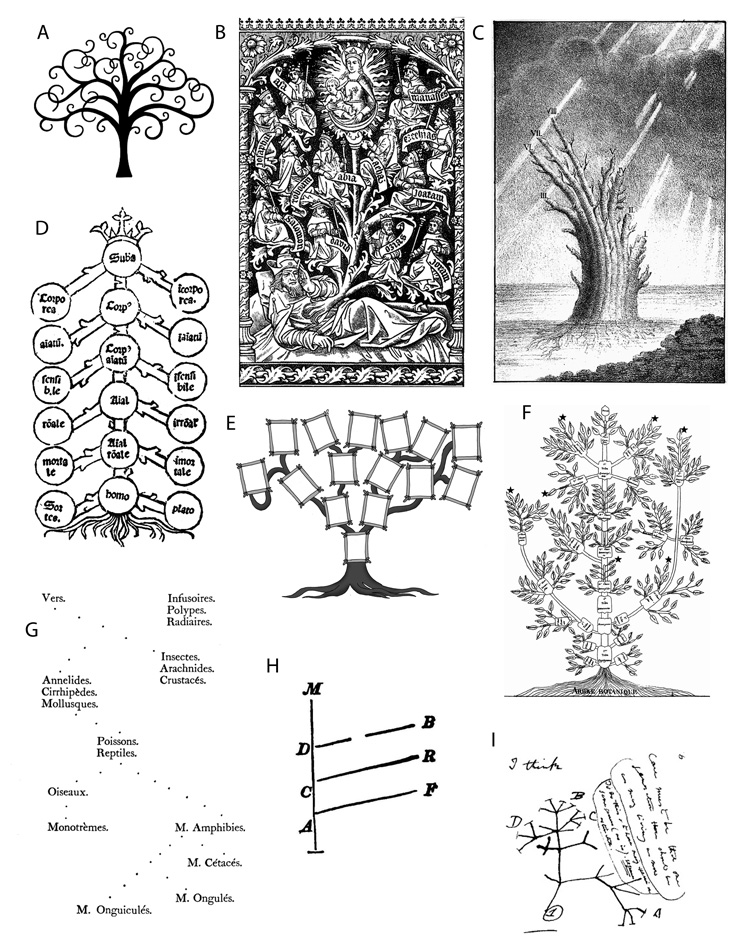
A fa mint az egyik leggyakoribb – és mindenképpen
a legfeltűnőbb – növényi növekedési forma már ősidők óta kiemelt
szerepet tölt be a különféle kultúrákban. Konkrét biológiai
jelentésétől eltérően, de attól teljesen sohasem elszakadva,
megjelenik mítoszokban, vallásokban, népmesékben, az irodalomban és
a képzőművészetben egyaránt (1/a
ábra). A vastag törzs, a fokozatosan vékonyodó ágak
sokasága, a gallyak és a rajtuk fejlődő levelek, virágok, termések
vagy éppen tobozok mindig is gyakori ihletői voltak az „Élet fája”
megjelenítőinek – az eredményt tekintve igen változatos tartalommal
és küllemmel.1 Legismertebb
talán a bibliai Édenkertben fejlődő élet fája és annak közvetlen
közelében a jó és a rossz tudásának fája. A benne rejlő szimbolikus
üzenetek mellett a fa hosszú ideje egy konkrétabb, de a növénytanhoz
már nem kötődő jelentéssel is bír: az elágazások rendszere alkalmat
ad emberek leszármazási, rokoni vagy éppen dinasztikus viszonyainak
bemutatására (1/e ábra). Az első közismert, és a 12.
századtól kezdődően gyakorivá lett ilyen jellegű ábrázolás
Jesszének, Dávid király apjának a „családfája”. Számos festmény,
fríz és templomi ablak örökíti meg az alvó Jesszéből eredő,
erőteljes, fává növő vesszőt, melynek ágain jelennek meg a későbbi
királyok, próféták, s maga Jézus is (1/b ábra). A zsidó
Lurie-család fája – melynek többek között Karl Marx, Sigmund Freud,
Felix Mendelssohn és Yehudi Menuhin is tagja – egészen Dávid
királyig, azaz több mint háromezer évvel ezelőttig vezeti vissza
történetüket (Rosenstein, 2004). A legnépesebb Konfuciusz összes
(ismert) utódjának a „családfája”, amely igen nagy időtávlatot,
nyolcvan generációt átívelve, s mintegy kétmillió nevet felsorolva
összesíti a nagy kínai gondolkodó leszármazottait (URL1).
Hasonló módon mutathatjuk be például Ádám és Éva utódainak rokonsági
viszonyait, királyi és császári uralkodóházak történetét vagy éppen
fiktív regényhősök családi kapcsolatait Harry Pottertől a dallasi
Ewing famíliáig.2 Hazai példa
is akad, elegendő, ha az Árpád-ház (URL3)
és a Rákócziak (URL4)
közismert „családfáit” felidézzük.
A fa fogalma a tudományban sem maradt meg eredeti
jelentésénél, melynek első jeleit – mint oly sok más esetben – már
az ókori görögök munkáiban is megfigyelhetjük. Arisztotelész
divíziós osztályozási alapelvei szerint a dolgok lényegük szerint
felső génuszokba, azokon belül speciesekbe sorolhatók, amelyek több
lépcsőben további génuszokra és speciesekre bonthatók, egészen a
legalsó fajokig. A divíziók sorozata hierarchiát alkot, ami fő
szerepet játszott az évszázadokkal később tevékenykedő, ugyancsak
görög filozófus, Porfiriusz egyik művében. Ennek latinba átültetett
kiadásai a génuszok/speciesek hierarchiáját már Porfiriusz fájaként
ismertették – és ábrázolták (1/d ábra). Egy ilyen absztrakt
fa szolgált alapul Carl von Linné számára az élővilág
klasszifikációjához, a taxonómiai rangok egymásba ágyazott
rendszeréhez.
A rendszerezésen kívül a fa hamarosan a biológiai
sokféleség metaforájává is vált. Az orosz cári udvarban évtizedekig
vendégeskedő német Peter Simon Pallasnak (1766) tulajdonítják sokan
azt a koncepciót, hogy az élő szervezetek közötti hasonlóságokat
(affinitásokat) fa formájában lehet bemutatni – bár ő maga sohasem
valósította ezt meg. A szintén német Carl Edward von Eichwald csak
hatvan évvel később vállalkozott erre egy misztikus környezetbe
helyezett fa-óriás megrajzolásával (1/c ábra). A francia
szerzetes-botanikus Augustin Augier is művészi ihletéssel készített
fával ábrázolta a növények osztályozását (Augier, 1801), melyben a
fa törzse a nagyobb csoportok tökéletlentől a tökéletesig való
elrendezését jelképezte, az ágak és a levelek pedig a kisebb
kategóriáknak feleltek meg (1/f ábra). Linné rendszere
azonban változatlanságot sugallt, és a hozzá eszmeileg kötődő
artisztikus faábrázolások sem voltak mások, mint az állandóság
szimbólumai.
A fajellegű struktúra, azaz elágazások összetett
rendszere sokkal inkább alkalmas az utódlás, a leszármazás vagy az
átalakulás bemutatására – ahogy azt a családfának nevezett
ábrázolásoknál már láttuk.3
Ez teljes egyértelműséggel először Jean-Baptiste Lamarcknál
jelentkezik a biológia történetében. Az állatok „transzmutációját”
egy – általa Tableau-nak nevezett, a különféle állatcsoportok
eredetére (l’origine) célzó – elágazó sémával (1/g ábra)
mutatta be 1809-ben (Lamarck, 1809, Podani, 2010, 1/c ábra).
Erre ma már nyugodtan ráragaszthatjuk az első „filogenetikai fa”
címkéjét. Annál is inkább, mert Lamarck kevés számú követőinek
egyike, Charles-Hélion de Barbançois ezt a fát nagy felbontásban,
poszter-méretben rajzolta meg 1816-ban, melynek aláírásában már
szerepelt az egyértelmű filiation (leszármazás) szó is. Egy
távolinak tűnő tudományterület, az embriológia is „bejelentkezett”:
az angol William Benjamin Carpenter a gerincesek egyedfejlődésének
korai szakaszai és a főbb gerinces csoportok között mutatkozó
hasonlóságokat ábrázolta egy fadiagrammal 1841-ben. Erre
kísértetiesen hasonlít a Charles Robert Darwint már közvetlenül
befolyásoló híres-hírhedt tudományos pamflet (Chambers, 1844) egyik
ábrája, amivel az anonimitását haláláig megőrző szerző az
embrionális fejlődés és az evolúció párhuzamosságát érzékeltette
(1/h ábra). S ahogy Pallas nem rajzolt, ugyanúgy megmaradt a
szavak szintjén az evolúciós gondolat két kiváló képviselője, a
francia Charles Victor Naudin és az angol Alfred Russel Wallace is.
Naudin egy 1852-es írása szerint a növények birodalma akár „egy fa
is lehetne, melynek a mélységes mély múltban rejtező gyökerei
néhány, egymás után kiágazó törzset fejlesztenek. Eme első törzsek
jelenthetik a birodalom legősibb formáit, míg az utolsó ágak a ma
élő fajoknak felelhetnek meg”. Wallace 1855-ös meglátása hasonló
volt: „a ma élő fajok prototípusai a hasonló szervezetek vonalainak
kusza szövedékét alkották, olyan finoman mint egy göcsörtös törzsű
tölgy gallyai vagy az emberi test véredényei”.
Ezen a ponton jogosan tehető fel a kérdés: hol a
helye Darwinnak ebben a történetben? Kortársai elképzeléseivel
természetesen tisztában volt, de közismert módon egész életében csak
egyetlenegy fadiagramot jelentetett meg, az On the Origin of Species
egyedüli ábrájaként (Darwin, 1859; Podani, 2010, 1d ábra),
amelyet „Az élet fájaként” aposztrofált. Ez korántsem jelenti azt,
hogy ne gondolkozott volna már jóval előtte, két évtizeden át arról,
mi is lehet az evolúció hiteles ábrázolása, legjobb grafikus
modellje. Az utókorra maradt jegyzeteiből kiderül, hogy már az
1830-as években több fajellegű ábrázolást is készített. Ezek közül
leghíresebbé az I think feliratot viselő rajz vált (1/i ábra),
ami tetoválásokon és trikókon is népszerűsíti Darwin elméletét
szerte a világon. (URL5
és
URL6 – ez utóbbinak darabja 20
USD fölött van.) Ezeken a diagramokon, a „Tree of Life”-ra való
utalások ellenére Darwin egyáltalán nem az egész élet leszármazási
kapcsolatait próbálta megmutatni, hanem a fajkeletkezés modelljét
érzékeltette, azaz jóval kisebb léptékben gondolkodott. Néhány
rajzán – ritka kivételként – egy konkrét rendszertani csoport,
nevezetesen a gerincesek rokonsági viszonyairól szóló elképzeléseit
is felvázolta.
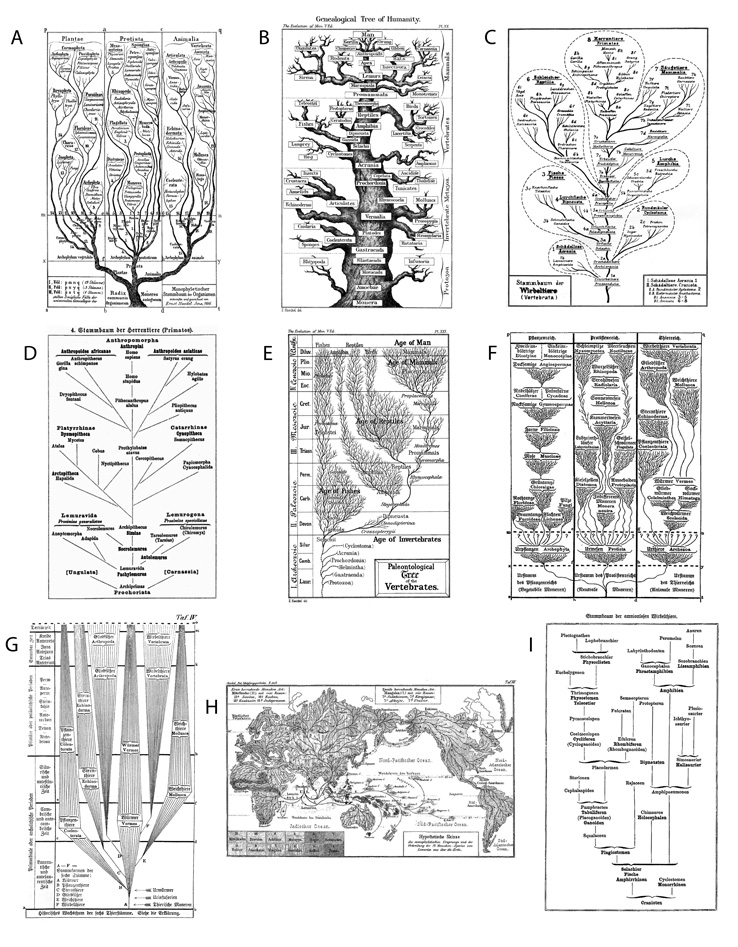
Az élet, vagyis a teljes akkor ismert élővilág –
leszármazási – fájának első ábrázolása Darwin tanainak
legszorgalmasabb európai terjesztőjére, Ernst Heinrich Philipp
August Haeckelre várt. A jénai professzor legalább akkora művész
volt, mint tudós, amit a Kunstformen der Natur (Breidbach, 2006)
csodálatos festményein kívül azzal is alátámaszthatunk, hogy az
evolúciós értelmezésű fadiagramok mindmáig párját ritkító
változatosságát, valóságos erdejét produkálta
(2. ábra). Kreativitását az is
igazolja, hogy neki köszönhető olyan fontos fogalmak bevezetése a
tudományba, mint a törzs (phylum), törzsfejlődés, törzsfa,
monofiletikusság, ökológia, protista vagy éppen az őssejt. Ezek egy
része szerepel annak az 1866-os ábrának az aláírásában is, amellyel
Haeckel az összes élő szervezet közös (azaz monofiletikus) eredetét
illusztrálta és – jelentős újításként – szakított a hagyományos
növény-állat felosztással, külön kezelve az egysejtűek (azaz a
protista) fő ágát (Monophyletische Stammbaum, 2/a ábra).
Később a botanikai fákra még inkább emlékeztető, bár erősen
stilizált fákat is készített, mint például az állatok „tölgyfája”4
(2/c ábra), melynek csúcsán – a klasszikus szemlélethez
hűen – az embert tüntette fel. Lamarckéhoz hasonló, vonalas-szöveges
diagramokon mutatta be minden sallang nélkül a növények, a főemlősök
(2/d ábra) vagy éppen a gerincesek (2/i ábra)
leszármazási viszonyait – ez utóbbiakon továbbra is a mi fajunkat
helyezve a legfelső pozícióba. A Hypotetische Skizze az ember
származását és szétterjedését illusztrálta egy térképre fektetett
kladogram-szerű rajzzal (2/h ábra). Haeckel a geológiai
időskálát is gyakran mellékelte, és a paleontológusok kedvenc
motívumát, a csoportok fajgazdagságával arányos vastagságú ágakat
alkalmazta (mint például az állatvilág esetében, 2/e és g
ábra).5 Érdekes kettősség
Haeckel grafikáin, hogy – míg az első Stammbaum az alá- és
fölérendeltséget sugalló arisztotelészi scala naturae (természet
lépcsője) hatásától mentes volt – rajzainak többsége mégis ilyen
szellemben készült, az evolúciós fokozatok, a morfológiai
„progresszió” érzékeltetésének szándékával.
Haeckelre nemcsak Darwin, hanem korának sok
nyelvésze is kimutatható hatással volt – már csak azért is, mert
személyesen ismerte néhányukat. Abban az időben teljesen egyértelmű
volt a párhuzam a biológiával: a nyelvek is változnak, nemcsak az
élő szervezetek. Erre Sir William Jones már 1786-ban célzott –
megjegyezve, hogy a szanszkrit, a görög és a latin nyelv egy közös,
ma már nem létező nyelvből vezethető le. Az igazi áttörésre két
fontos közlemény 1853-as, egymástól – valószínűleg – független
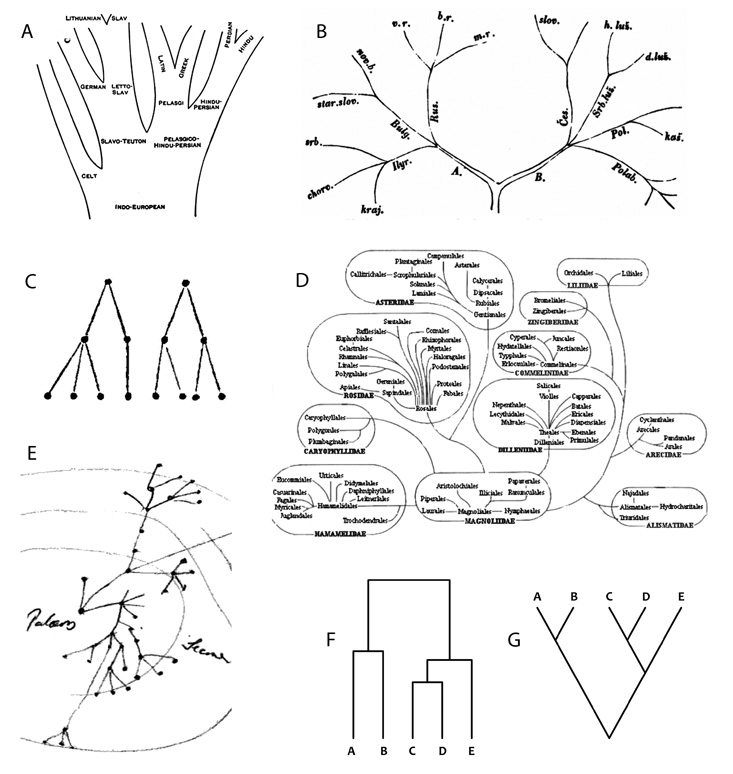
megjelenéséig kellett várni. A jénai August Schleicher az
indoeurópai nyelvek stilizált fáját vázolta fel, a cseh František
Ladislav Čelakovský posztumusz megjelent rajza pedig a szláv
nyelvekre összpontosított. Bár egy ideig kollégák is voltak a prágai
egyetemen, és Schleicher csehül is publikált, semmi bizonyíték nincs
arra, hogy összebeszéltek vagy plagizáltak volna – amit a két rajz
igen eltérő grafikai megvalósítása is igazolhat
(3/a–b ábra) (Priestly, 1975).
A nyelvek és az élő szervezetek evolúciójának elvi hasonlatosságait
egyébként maga Darwin is több helyen fejtegette az Origin-ben. A
nyelvrokonság „családfás” bemutatása mindennapos gyakorlattá vált a
lingvisztikában – és a nyelvek és a népek fájának statisztikai
összevetésére is van példa (Penny et al., 1993). A teljesség
kedvéért azt is megjegyezzük, hogy az evolúció „képessége” nincs a
szervezetekre és a nyelvekre korlátozva, így a fa hatékony
segédeszköz lehet kéziratokban és egy adott könyv különböző
kiadásaiban fellelhető változások bemutatásában, vagy éppen zenei
stílusok és irányzatok összefüggéseinek érzékeltetésében is.
A fa jelentése tehát sokféleképpen módosult a
tudományban, és némi meglepetéssel vehetjük észre, hogy a matematika
kissé lemaradt a logika, a biológia és a nyelvészet mögött. Az angol
Arthur Cayley 1857-ben pótolta ezt a „hiányosságot”: a fát
gráfelméleti objektumként definiálta, amely szögpontok és az azokat
összekötő élek halmazából áll, azzal a megkötéssel, hogy egy adott
szögpontból bármely másikba csak egyféleképpen lehet az élek mentén
eljutni (3/c ábra) (Cayley, 1857). Így pontosította azt az
alapvető „fasajátságot”, miszerint ami már egyszer elvált egymástól,
az tovább ágazhat ugyan, de sohasem egyesülhet újra – ellentétben
például a levelek anasztomizáló erezetével.6
Más szóval, a fában nincsenek körök. Figyelemre méltó tény, hogy
Darwin jegyzetkönyveiben is találunk olyan rajzokat, amelyek
tökéletesen megfelelnek Cayley meghatározásának, azaz egy
gráfelméleti fának. 1857-ből, vagy egy évvel későbbről származik a
halak leszármazását illusztráló ábra, melyen a mellette található
jegyzetek szerint a pontok evolúciósan „új formáknak” felelnek meg
(3/d ábra). A koncentrikus körökkel pedig Darwin az eltelt
időszakokat kívánta szemléltetni. Az időbeli egybeesés ellenére
nagyon valószínű, hogy Darwin nem olvasta Cayley cikkét, holott
ismerhette őt a Royal Societyből.7
Darwin és Haeckel nyomán a fahasonlat igen
népszerűvé vált mind az állattanban (az angol William Abbott Herdman
könyvében), mind a növénytanban (az amerikai Charles Edwin Bessey,
és a német Hans Hallier munkáiban). A 20. század közepe táján
elsősorban a botanikusok alkalmazták előszeretettel – nem
változtatva azon a szokáson, hogy először készült el az osztályozás,
amire utóbb próbálták az egyes csoportok – feltételezett –
filogenetikai viszonyait fa formájában vetíteni. Az ilyen stratégiát
követő, úgynevezett filogenetikai rendszerezők közé tartozik például
az örmény Armen Tahtadzsján (Takhtadjan), az amerikai Arthur
Cronquist (3/e ábra) és a magyar Soó Rezső is.
A sorrend megváltoztatása – vagyis készüljön
először egy fa, majd utána próbáljunk meg annak alapján osztályozni
– két, a matematikai módszereket és a számítógépet intenzíven
alkalmazó rendszertani irányzatra, a numerikus taxonómiára és a
kladisztikára várt. Az amerikai rovarász, Robert Reuven Sokal és az
angol mikrobiológus, Peter Henry Andrews Sneath által népszerűsített
és matematikusok által továbbfejlesztett numerikus taxonómiai
módszerek előnye a viszonylagos objektivitás volt. Elsősorban
morfológiai adatok alapján számítógép állított elő egy fadiagramot
(dendrogramot, 3/f ábra), ami a vizsgált szervezetek
hasonlóságainak összegzője: minél hasonlóbb két szervezet vagy
szervezetek két csoportja, annál előbb kapcsolódnak egymáshoz. Az
ábrán C és D a leghasonlóbbak, majd az E következik. Az A és B
valamivel kisebb hasonlóságot mutat fel, és egyértelműen külön
csoportot alkot. A kutató a dendrogram alapján határolta el a
taxonokat – ezt már kevésbé objektív módon. Publikált dendrogramok
ezrei jelezték az irányzat népszerűségét, és a gráfelméletileg is
tiszta jelentésű fafogalom meghonosodását a biológiában. A numerikus
taxonómusok abban tévedtek, hogy az evolúció részleteiben sem
rekonstruálható, s emiatt természetes osztályozást csak a
szervezetek külső hasonlósága alapján tudtak elképzelni. A német
Emil Hans Willi Hennig azonban őket pár évvel megelőzve már
kidolgozta a kladisztika elméleti alapjait, s megmutatta, hogyan
lehet a jelen ismeretében a múltra következtetni. Ezzel
megvalósította Darwin javaslatát, miszerint a leginkább természetes
klasszifikáció a leszármazáson alapszik – tehát ez utóbbit kell
először megismernünk. A numerikus taxonómia valójában csak kikövezte
az utat a számítógépre egyre inkább hagyatkozó kladisztika számára,
ami az 1980-as évektől fokozatosan átvette a vezető szerepet a
szisztematikában, és a biológiai kutatások nélkülözhetetlen
összetevőjévé vált. A kladisztika paradigmatikus ábráján, a
kladogramon (3/g ábra) nem a hasonlóság, hanem a
filogenetikai rokonság fejeződik ki. Az ábrán az A és a C, valamint
a C és a D egymás legközelebbi rokonai, míg az E az utóbbiak kissé
távolabbi rokona. Morfológiai, de főként molekuláris alapon végzett
elemzések sokasága kladogramok tízezreit eredményezte és
szolgáltatja ma is, amivel egyre közelebb jutunk a nagy álom
megvalósításához, az Élet „fájának” (pontosabban kladogramjának, vö.
Podani, 2010) megszerkesztéséhez. A „fában gondolkodás”
(„tree-thinking”), ahogy Robert O’Hara 1988-ban megfogalmazta, a
biológiai érvelés és gondolkodás nélkülözhetetlen részévé vált, s
nélküle nem képzelhető el az evolúciós folyamatok elemzése,
bemutatása, magyarázata és megértése sem. Egy kérdés azonban
továbbra is nyitott: mindenki ugyanabban a fában gondolkodik-e, s ha
igen, vajon ugyanolyan módon? Minden rendben van-e a fában
gondolkodással? Elősegítette-e a fa matematikai definíciója a
fadiagramok egyértelmű rendszertani alkalmazását? A válasz sajnos
nemleges, ami hamar kiderül azon huszonegy tudományos cikk és egy
könyv elolvasásából, melyek már a címükben is feltüntetik a
tree-thinking kifejezést. A problémák egy részére már céloztam
(Podani, 2010) – de maradt bőven megbeszélni való.
Családfák – nem mindig család, és nem mindig fa
Az olvasónak feltűnhetett, hogy a családfa szó több helyen is
idézőjelben szerepelt. Ezzel arra utaltam, hogy a fán szereplő
személyek köre általában lényegesen túllépi a család megszokott
kereteit, azaz a „vér szerinti rokonok közösségét, melynek tagjai
együtt élnek” (URL8).
Nyilván senki sem gondolja komolyan, hogy Jessze vagy éppen
Konfuciusz összes leszármazottja egy családba tartozik – még a
legtágabb értelemben sem. A „család” előtag tehát egy túlzó
általánosításnak, félreértésnek köszönhetően tapadt szinte
kitörölhetetlenül a „fá”-ra, nem csak a mi nyelvünkben. Családfáról
még kevésbé beszélhetünk, amikor csak egy adott személy (általában
egy férfi) leszármazottjai, vagyis gyermekei, unokái, dédunokái
szerepelnek rajta – hiszen egy család nem család a házastársak
nélkül. Gyakran így nevezik a „fordított fát” is, melynek levelei a
legrégebbi ősök, s tőlük kiindulva a fa törzse felé haladva jutunk
el az ükapákon, dédnagymamákon, majd a szülőkön keresztül a fa
tövéig, egyetlen személyig, akinek tehát az összes felmenője – és
nem utódja – adja a fa ágrendszerét és lombozatát. De erre az
ábrázolásra legalább van egy másik, francia eredetű szavunk, a
pedigré (pied de grue – daruláb). A helyzet bonyolultságát jelzi,
hogy amikor a feltüntetendő személyek tényleges családot, de
legalábbis egy kellően szűk rokonsági kört képeznek, akkor a
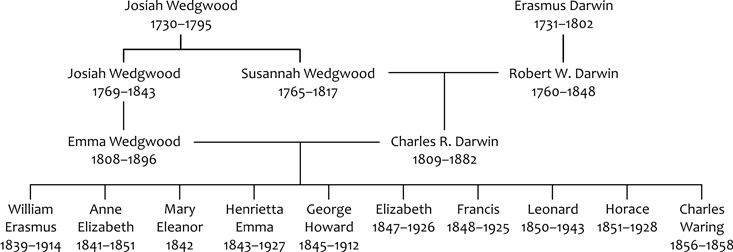
fahasonlat válhat helytelenné. A legjobb példa Darwin „családfája”
(4. ábra), amelyen a
házastársak is szerepelnek, viszont Darwin felesége, Emma, az
unokatestvére volt – a gráfban tehát bezárul egy kör, s fáról
definíciószerűen nem beszélhetünk. Itt tehát már nem – az általam
igen amatőr módon művelt – nyelvészkedésről van szó csupán, hanem a
matematikai fasajátság megsértéséről. S miután Konfuciusz milliónyi
utódja között is biztos voltak igen közeli rokonságban állók,
mondjuk házastársként, se a „család” se a „fa” nem tükrözi
megfelelően a tartalmat – s valójában emberek leszármazási
hálózatáról (genealogical network) kellene beszélnünk.
Tudjuk persze, hogy a család szó több jelentéssel
bír. A biológiában, például a Linné-féle hierarchiában az egyik
rangnak felel meg, a legfontosabbnak a génusz és a rend között. Az
igazsághoz tartozik ugyanakkor, hogy maga Linné nem használta a
család (familia) kategóriáját, noha azt a francia Pierre Magnol
(1689) már jóval a nagy svéd természettudós megszületése előtt
alkalmazta – amikor viszont a rangok hierarchiája volt
kidolgozatlan. Philosophia botanica című, alapvető fontosságú
művében Linné a familia szót a növények birodalmának (Vegetabilia)
egy nem rendszertani célú csoportosításában használta csupán.8
Csak a 19. században rögzült a család rangja a rendszertanban.9
Rendszertani nézőpontból a családfa elnevezés tehát legfeljebb akkor
lenne elfogadható, ha ténylegesen egy család, mondjuk az Asteraceae
(fészkesek) vagy a Felidae (macskafélék) fajainak leszármazási
mintázatát ábrázolnánk a fadiagrammal.
A Haeckel által bevezetett törzsfa fogalom is
hasonló problémákkal terhelt. Miféle törzsnek milyen fájáról is
beszélünk? – kérdezhetjük. A törzs (Stamm) szó egyik jelentése
valóban lefed egy közös eredetű csoportot (mint például bennszülött |
|
|
törzsek vagy baktériumtörzs) de emellett
rendszertani kategória is, hasonlóan a családhoz. Talán már eleve a
német stammen szóra kellett volna gondolni, melynek jelentése többek
között származni, eredni, tehát leszármazási fának fordítani Hackel
Stammbaum-ját. A latinizált változat, vagyis a phylum sem javít
sokat a helyzeten, lévén az is rendszertani kategória egyben – de
idegen szóként talán jobban befogadható, s filogenezis, illetve
filogenetika alakjában jobban beilleszkedhet a nyelvünkbe.
A következő felsorolás megmutatja, hogy egyes
élőlénycsoportok leszármazását bemutató fadiagramokra hányféleképpen
és mekkora redundanciával vagy pontatlansággal hivatkoznak manapság
mind a magyar, mind az angol nyelvterületen: evolúciós családfa,
filogenetikai törzsfa, filogenetikus családfa (rokonsági fa),
törzsfa (dendrogram), illetve phylogenetic family tree, evolutionary
family tree, evolutionary phylogenetic tree, phylogeny (cladogram)
és phylogenetic evolutionary tree – azaz számos kombinációban.
Forrást nem jelöltem meg egyik esetben sem, mert igazságtalan volna
bármelyik szerzőt vagy fordítót kiemelni az inkriminált cikkek és
könyvek sokaságából. Az olvasó persze a fenti érvrendszert nyelvi
okoskodásnak, sőt akadékoskodásnak vélheti, mondván: mindezzel
legfeljebb a dolgok felszínét érintettük. Valóban, vannak itt
lényegesebb problémák is. A megnevezések és a fák megjelenítésének
és értelmezésének módozatai azonban nem fednek át egymással –
ugyanazon jelentésű fára többféle néven is hivatkoznak, illetve egy
konkrét fogalmat eltérően értelmezendő fákra is alkalmaznak. Többről
van tehát szó, mint redundanciáról vagy szinonima-túltengésről –
hiszen valóságos terminológiai káosszal állunk szemben. A
szakzsargon pontosítására a következő részben teszek majd
javaslatokat. Egy fogalmat azonban már előre kizárhatunk a szóba
jövő lehetőségek köréből. A dendrogram egyértelműen NEM evolúciós
jelentésű fadiagram, hanem általában a (hierarchikus) cluster
analysis eredményeinek ábrázolási formája. E módszerek csak egyik,
bár tudománytörténeti szempontból fontos alkalmazási területe a
numerikus taxonómia. A dendrogram filogenetikai interpretációja
viszont nehézségekbe ütközik, még akkor is, ha molekuláris adatokból
állítjuk elő – s ma már nem is alkalmazzák filogenetikai
kutatásokban.
Az idő és a szervezetek közötti relációk kérdése
A fadiagramok nem csak ránézésre, grafikus megjelenésükben
különböznek egymástól. Még nagyobbak az elvi eltérések, azaz
miképpen viszonyulnak az idődimenzióhoz, és milyen formában fejezik
ki az evolúciós rokonságot. Természetesen számunkra ezek a
legfontosabb, és tovább részletezendő szempontok. Az első
filogenetikus fa, Lamarcké (1/g ábra), egymagában példáz két
lehetőséget is. A hüllők (Reptiles) és a madarak (Oiseaux) között
közvetlen az összeköttetés csakúgy, mint a puhatestűek (Mollusques)
és a halak (Poissons) között. A patás emlősök (M. ongulés) és a
karmosak (M. onguiculés) viszont már másféleképpen, egy belső
szögponton keresztül kapcsolódnak csak egymáshoz. Ugyanez a viszony
áll fenn a férgek (Vers.) utáni két hármas csoport között is.
Vizsgáljuk most meg alaposan ezt az ábrázolásbeli kettősséget friss
szemmel, több mint kétszáz évvel az eredeti publikáció után!
A ma is élő fajokat magába foglaló rendszertani
kategóriák összekötésével a francia természettudós nem csak azt
fejezte ki, hogy a madarak a hüllőktől származnak (értsd: az első
madárnak tekinthető élőlény egy, még hüllőnek tekinthető állat
leszármazottja volt). Munkásságának ismeretében elmondhatjuk, inkább
azt akarta jelezni, hogy a hüllő állapothoz képest a madarak
egyfajta progressziót (azaz „fejlettebb szintet”) képviselnek.
Lamarck ugyanis a több ezer éves rendező elv, a scala naturae
hatásától még nem tudott megszabadulni (éppen elég nagy fegyvertény
volt részéről a változatlanság hittételének cáfolata). Ebben a
természeti objektumok fokozatokat, a legalacsonyabb rendűnek
tekintett víztől, tűztől, kőzetektől a növényeken, majd az állatokon
át az emberig, sőt egyes ábrázolásokon még tovább, a
természetfelettiekig vezető sorozatot alkottak. Mint már említettük,
Haeckel ábráinak döntő többsége is a scala naturae hatása alatt
készült. Haeckel tehát nem a darwini, hanem a lamarcki, sőt az
arisztotelészi örökséget tartotta életben, s ültette át evolúciós
kontextusba. Az egymásutániság dogmájának hatása érvényesült az
amerikai Bessey növényosztályozásában, és a 20. század első
kétharmadának filogenetikai rendszereiben is. A kiindulópont mindig
valamilyen fajfeletti osztályozás volt, majd csak ezután következett
az evolúciós interpretáció a taxonok fába illesztésével. Ezeken a
fákon legalul, középen és legfelül is olyan csoportok vannak,
amelyek ma is élő fajokat foglalnak magukba, tehát a fa nincs az
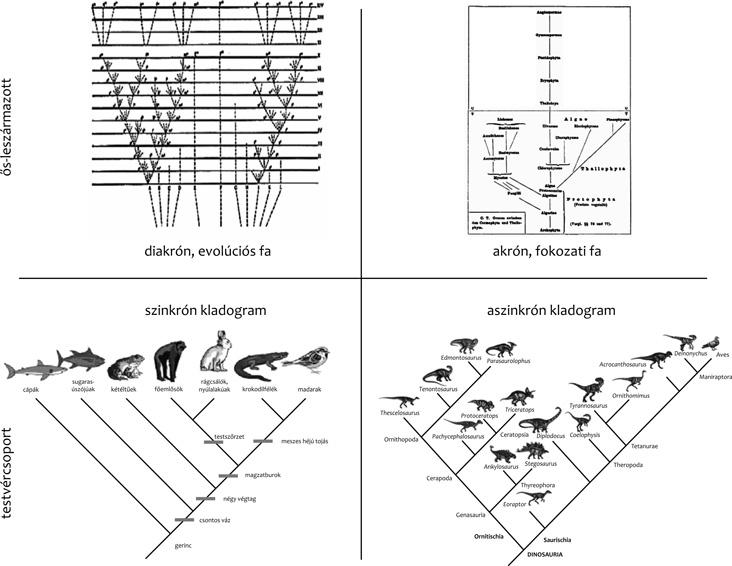
idődimenzióba beágyazva. Más szóval, akrón (gör. idő nélküli)
fokozati fákról beszélhetünk (achronous vagy grade trees, Podani
2013).
A patás és karmos emlősök Lamarck feltételezése
szerint nem egymásból alakultak ki, hanem két egyenrangú fokozatot
képviselnek, és valamely régen kihalt, ma már nem létező
ujj-szerkezetű emlőstípus kései utódai. Közös eredetük a fán úgy
mutatkozik meg, hogy egy belső szögponton vagy akár „hipotetikus
közös ősön” keresztül kapcsolódnak egymáshoz. Ezzel Lamarck –
feltehetően nem teljesen szándékosan – a patások és karmosok
testvércsoporti10
viszonyát mutatta be, a kladogramok egyfajta előképeként. Haeckel
néhány fája is így értelmezhető, különösképpen a legelső Stammbaum
(2/a ábra). Minden megnevezett csoport recens és a legvégső
gallyakon helyezkedik el – ősök és kihaltak nem szerepelnek a
rajzon.11 A név nélküli
belső szögpontok később is felbukkannak például Herdman és Hallier
fáin, valamint – akrón részekkel kombinálva – a növények
filogenetikai rendszereiben, mint például Tahtadzsjannál és
Cronquistnál (3/e ábra). Ez a koncepció a Darwin-féle
javaslatokat (vö. Podani, 2010) leginkább komolyan vevő Hennignél, a
kladisztika megalapítójánál kristályosodik ki teljesen. Hennig fája,
a kladogram, a testvércsoportok teljes rendszere, azaz lehetőség
szerint minden belső szögponton villás (dichotomikus) elágazású. Az
elemzésbe bevont szervezetek mindegyike egy időben, vagyis ma él, és
a fán végső helyzetben van. A szinkrón (gör. egyidejű) kladogram
elnevezés különbözteti meg a többi evolúciós interpretációjú
fadiagramtól („synchronous trees”, Podani 2013). Ha meggondoljuk, az
alapötlet zseniálisan egyszerű: a leszármazási mintázatot a ma élők
testvércsoporti relációival, és nem az ős–leszármazott kapcsolatok
bemutatásával is érzékeltetni tudjuk. Ráadásul a kladogram váza
mindhárom fajkeletkezési modell (hasadás, lefűződés és anagenezis12)
esetében is tükrözi a folyamat lényegét.
Az ideális persze az lenne, ha az ős–leszármazott
viszonyokat az eltelt idő függvényében is be tudnánk mutatni. A
leszármazás pontosan csak a fajok vagy populációk szintjére
ereszkedve érzékeltethető, a „hüllőből lett a madár” és hasonló
állítások erre alkalmatlanok – vagyis meg kellene szabadulnunk a
fokozati fákban alkalmazott magasabb rangoktól. Az ősöket azonban
nem ismerjük, és ha egy csoportban viszonylag sok is a fosszilis
lelet, akkor se tudunk megbizonyosodni arról, hogy azok ténylegesen
a maiak ősei vagy oldalági rokonai voltak. Számos nagy csoportnál
pedig a paleontológiai adatok hiánya vagy esetlegessége miatt ez a
kérdés eleve fel sem merül. Az Origin of Species ábrája és Darwin
sok más rajza tehát egy elméleti modell, a valós leszármazási
viszonyok csak legfeljebb rövid időtávlatban, például vírusoknál
vagy laboratóriumban lejátszott evolúciós kísérleteknél ismertek. Az
idődimenzióba ágyazott, ős–leszármazott viszonyokat bemutató ábrákat
a diakrón (gör. időbeli) vagy evolúciós fa elnevezéssel
különíthetjük el a többiektől.
Az időbeliség figyelembe vétele vagy mellőzése,
illetve az ős–leszármazott vagy testvércsoporti relációk bemutatása
két egymástól független alternatíva. Három kombinációt már láttunk,
lennie kell tehát egy negyedik lehetőségnek is, amelyben
testvércsoporti relációk mutatkoznak oly módon, hogy nem vagyunk
tekintettel arra, mikor is éltek a vizsgált szervezetek. Ennek
megfelelő fadiagram adódik, ha ma élő szervezeteket és kihalt
rokonaikat egyszerre vetjük kladisztikai elemzés alá
(értelemszerűen: legfeljebb morfológiai alapon,
5. ábra). Vigyázat, e fák
értelmezésével óvatosnak kell lennünk, hiszen egy kihalt és egy ma
élő faj testvércsoporti kapcsolata hamis lehet, ha az előző
tényleges őse volt az utóbbinak. Mint fentebb már céloztunk rá,
sohasem fogjuk megismerni, mi volt az igazság. Mintázati
kladisztikának (pattern cladistics) nevezik azt az irányzatot,
amelyet ez egyáltalán nem zavar, mert nem az elágazások rendszerét,
az evolúciós utakat akarja feltárni, csak a kládok tartalmára kíván
következtetni. Megelégszik azzal, ha a rokonság bármely formáját
(akár testvér, akár ős) ki tudja mutatni. Mivel a végágakon sérül az
egyidejűség feltétele, az ilyen fákat aszinkrón (nem egyidejű)
kladogramoknak („asynchronous trees”) (Podani 2013) nevezhetjük. A
múlt és a jelen szervezeteinek együttes, a leszármazásra épülő
osztályozására csak ilyen kladogramok alkalmasak – tehát a
rendszertan nélkülözhetetlen eszközei.
Fák vagy korallok?
Tankönyvekben és szakcikkekben az evolúciós folyamatok ismertetése
általában a populációk szintjén történik, az egyedek közötti
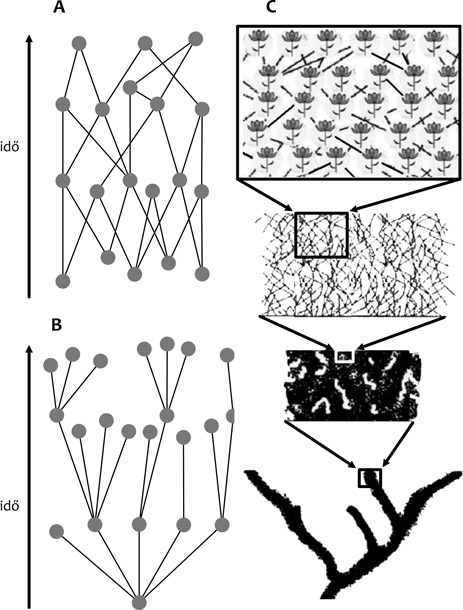
szülő–utód kapcsolatok bemutatásával. Egy ivaros szaporodású
populációra a megfelelő illusztráció egy hálózat, míg ivartalan
szaporodás esetében fa (6/a–b
ábra), amelyeket Hennig nyomán tokogenetikus gráfnak
nevezünk (gör. tókosz – kb. szülés, születés). A speciációt, vagyis
a populációk szétválásának modelljét nagy időtávlatba helyezve
azonban ez a megoldás nem működik. A lépték fokozatos növelésével a
grafikus részletek ugyanis „összefolynak”, és a diagram egyre inkább
emlékeztet tényleges biológiai fákra, helyesebben azok árnyképeire
(6/c ábra). Az ilyen ábrák aláírásában jó esetben csak a
filogenezis (phylogeny) szó szerepel, de gyakori a „filogenetikai
fa” megjelölés is. Matematikai szemszögből persze ezek egyáltalán
nem fák, hiszen nincsenek bennük élek és szögpontok. Kladogramnak
tűnő vonalrajzos diagramok pedig még inkább becsaphatnak bennünket,
ha a szerző a fa éleit tekinti fajoknak, és a szögpontokhoz
fajképződési eseményeket rendel (ún. stem trees) – ami nem mindig
nyilvánvaló a kísérő szövegből sem. Ez azért is problematikus, mert
a terminális élek ekkor a „semmibe” mutatnak: a végső szögpontok nem
azonosíthatók fajkeletkezési eseményként – vagyis a diagram
jóindulattal sem tekinthető fagráfnak. A sziluettek és azok
legkarcsúbb változatai egy egészen más jellegű ábrázolási módot
képviselnek. Értelmezésükben ismét az evolúciós elmélet egyik
atyjáig kell visszamenni a biológia történetében.
Darwin egyes rajzai, mivel nem adta meg
szabatosan (ahogy ma elvárnánk) az élek és a szögpontok jelentését,
nem teljesen egyértelműek. A közismert I think feliratot viselő fa
(1/i ábra) például – mai szemmel nézve – nemcsak gráfelméleti
fának, hanem stem tree-nek is felfogható. A jegyzetfüzet
elolvasásából kiderül, hogy inkább az utóbbi a helyes interpretáció.
Viszont, korai gondolatait papírra vetve Darwin nem is fában, hanem
korallokban gondolkodott. 1837-ben vagy ’38-ban írhatta (Darwin,
1837–1838) a következő, ma már egyre gyakrabban idézett mondatát:
„The tree of life should perhaps be called the coral of life,base of
branches dead; so that passages cannot be seen.”13
Darwin sokat foglalkozott korallokkal, és jól
tudta, hogy számos fajuk gazdagon elágazik, és az ágak legfeljebb
alul olvadnak össze.14 A
korall hasonlat ráadásul még jobb is a fáénál, hiszen a korallok
alsó, idősebb részei fokozatosan elhalnak, akár az elmúlt korok
élőlényei. A fa viszont, ha vannak is benne üres sejtek, a
legvékonyabb gyökerektől a levelekig teljes egészében élő – tehát
csak elágazó szerkezetében utalhat a filogenezisre! Továbbá, a fa
törzse jóval vastagabb az ágaknál és gallyaknál, azt sugallva, hogy
a múltban több egyed élt, vagy nagyobb volt a biodiverzitás, mint a
jelenben. A végig nagyjából egyenletesen vastag, itt-ott kidudorodó
korall tehát jobban jelképezheti az egyedszám és a biodiverzitás
arányait, mint a felfelé vékonyodó faágak. Később azonban, talán a
szüntelenül fákat emlegető kortársak hatására Darwin elállt ettől az
ötlettől, s maga is következetesen a fa metaforáját alkalmazta. A
phylogeny és stem tree jellegű ábrázolások viszont inkább éppen a
korallnak felelnek meg: minden egyes pontjuk felfogható egy valaha
élt, vagy éppen most is élő szervezetnek, akár a korallokat alkotó
kis polipok. Célszerű lenne tehát „feléleszteni” Darwin fiatalkori
ötletét, és a nem gráf jellegű elágazó sziluettekre inkább a korall
elnevezést alkalmazni. Ha a fa fogalma precíz meghatározást
nyerhetett a matematikában, akkor a korall módosult jelentése is
rögzülhet előbb-utóbb a tudományos nyelvezetben. Ez fontos lépés
lenne ahhoz, hogy a „fában gondolkodás” ne csak egy szlogen
maradjon, hanem a tartalmában is egyértelműen elkülönüljön a
„korallban gondolkodástól”.
Záró megjegyzések
Az evolúció ábrázolásában képileg és tartalmilag is komoly a
zűrzavar, melynek történeti háttere a fa fogalmának sokfélesége, a
mitológia szimbolizmusától a szabatos gráfelméleti meghatározásig.
Biológiai értelemben döntő, hogy egy fajellegű ábrázolás figyelembe
veszi-e vagy mellőzi az időbeliséget, illetve az evolúció folyamatát
közvetlen ős–leszármaztatott vagy a testvércsoporti relációk
bemutatásával próbálja-e kifejezni. Ezek a szempontok a
filogenetikai értelmezésű fagráfok négyes csoportosítását
eredményezik. Az irodalomban és a köznapi nyelvben alkalmazott
szakzsargon meglehetősen zavaros, és félreértésekre vezethet, ami az
általam javasolt terminológia révén kiküszöbölhető. A klasszikus,
természet lépcsőjéhez kötődő gondolkodás maradványai, a fokozati fák
a nagyléptékű változások összesítői. A legfinomabb léptékben az
evolúciós fák működnek, de elsősorban csak modellekben, például
oktatási céllal. A múlt „rekonstrukciójára” csak kladogramok
alkalmasak: ha csak a jelen élővilágára összpontosítunk, akkor az
evolúciós folyamatok és azok eredménye is megjeleníthető, a múlt
szervezeteinek bevonásával már csak egy általános rokonsági reláció
elemezhető – amely azonban nélkülözhetetlen a mindenkori élet
osztályozásában, a történetiség értelmezésében.
A kézirathoz fűzött hasznos megjegyzéseiért köszönetemet fejezem ki
Kontra Klárának, Rózsa Lajosnak, Simoncsics Péternek és Szilágyi
Andrásnak.
Kulcsszavak: törzsfa, dendrogram, hierarchia, korall,
leszármazás, dichotómia, testvércsoport, aszinkrón
IRODALOM
Augier, Augustin (1801): Essai d’une
nouvelle classification des végétaux. Lyon: Bruyset Ainé et Comp.
•
WEBCÍM
Archibald, J. David (2014): Aristotle’s
Ladder, Darwin’s Tree. New York: Columbia University Press
Baum, David A. – Smith, Stacey D.
(2013): Tree Thinking. Greenwood, CO: Roberts and Co.
Breidbach, Olaf (2006): Visions of
Nature: The Art and Science of Ernst Haeckel. München: Prestel
Verlag
Cayley, Arthur (1857): On the theory of
the analytical forms called trees. Philosophical Magazine 4th Series
13, 85, 172–176.
Chambers, Robert (1844): Vestiges of the
Natural History of Creation, London: John Churchill •
WEBCÍM
Darwin, Charles (1837–1838) Notebook B.
•
WEBCÍM
Darwin, Charles (1842): The Structure
and Distribution of Coral Reefs. Being the First Part of the Geology
of the Voyage of the Beagle, under the Command of Capt. Fitzroy, R.
N. during the Years 1832 to 1836. London: Smith Elder and Co. •
WEBCÍM
Darwin, Charles (1859): On the Origin of
Species. London: Murray •
WEBCÍM
Dayrat, Benoît (2003): The Roots of
Phylogeny: How Did Haeckel Build His Trees? Systematic Biology. 52,
515–527. doi:10.1080/1063515039021827 •
WEBCÍM
Hull, David L. (1985): Darwinism as a
Historical Entity: A Historiographic Proposal. In: Kohn, David (ed.)
The Darwinian Heritage. Princeton, NJ: Princeton University Press,
773–812.
Kutschera, Ulrich (2011): From the Scala
Naturae to the Symbiogenetic and Dynamic Tree of Life. Biology
Direct. 6, 33. DOI: 10.1186/1745-6150-6-33 •
WEBCÍM
Lamarck, Jean-Baptiste (1809):
Philosophie zoologique. Paris: Dentu •
WEBCÍM
Magnol, Pierre (1689): Prodromus
historiae generalis plantarum, in quo familiae plantarum per tabulas
disponuntur. Montpellier •
WEBCÍM
Penny, David – Watson, Elizabeth E. –
Steel, Michael A. (1993): Trees from Languages and Genes Are Very
Similar. Systematic Biology. 42, 3, 382–384. DOI:
10.1093/sysbio/42.3.382
Pietsch, Theodore W. (2012): Trees of
Life. A Visual History of Evolution. Baltimore, MD: John Hopkins
University Press
Podani János (2010): Evolúció, törzsfa,
osztályozás. Magyar Tudomány. 171, 10, 1179–1192. •
WEBCÍM
Podani János (2013): Tree Thinking, Time
and Topology: Comments on the Interpretation of Tree Diagrams in
Evolutionary/Phylogenetic Systematics. Cladistics. 29, 315–327.
DOI: 10.1111/j.1096-0031. 2012.00423.x •
WEBCÍM
Podani János (2016): A növények
evolúciója és osztályozása. Rendhagyó rendszertan. Budapest: Eötvös
Kiadó
Priestly, Tom M. S. (1975): Schleicher,
Celakovsky, and the Family-Tree Diagram. Historiographia
Linguistica. 2, 299–333. DOI: 10.1075/hl.2.3.03pri
Rosenstein, Neil (2004): The Lurie
Legacy. The House of Davidic Royal Descent. Bergenfield, NJ:
Avotaynu Tartalomjegyzék •
WEBCÍM
Tassy, Pascal (2011): Trees before and
after Darwin. J. Zoological Systematics and Evolutionary Research.
49, 89–101. DOI: 10.1111/j.1439-0469.2010.00585.x
Wiley, Edward O. – Lieberman, Bruce S.
(2011): Phylogenetics. Theory and Practice of Phylogenetic
Systematics. Hoboken, NJ: Wiley and Blackwell DOI: 10.1002/
9781118017883.ch8
LÁBJEGYZETEK
1 Erről könnyen
meggyőződhetünk, ha a Google képkeresőjébe beírjuk a tree of life
kifejezést. <
2 Meglehetősen kimerítő
lista:
WEBCÍM
<
3 Bár a szervezetek
megváltozásának, transzmutációjának gondolata grafikusan először
tulajdonképpen hálózatok és nem fák formájában jelentkezett,
mégpedig Georges-Louis Leclerc, Buffon grófja kutyafajtákat és
Antoine Nicolas Duchesne eperfajtákat bemutató rajzain 1755-ben,
illetve 1766-ban.
<
4 Erre a rajzra a
legtöbb forrás Haeckel’s oak néven hivatkozik.
<
5 Az ilyen jelentésű
ábrák később orsó-, buborék- vagy Romer-diagram néven terjedtek el,
ez utóbbi az amerikai paleontológus, Alfred Sherwood Romer huszadik
századi aktivitásának köszönhetően.
<
6 Szabályt „erősítő”
kivételek persze vannak, hiszen ágak és törzsek összeolvadása is
megfigyelhető a természetben (például a Ficus-nál).
<
7 Tudjuk, persze, hogy
Darwin meglehetősen visszavonultan dolgozott downe-i házában, s elég
ritka vendég volt a tudományos konferenciákon s egyéb
összejöveteleken. A Darwin Correspondence Project (WEBCÍM)
szerint sohasem váltottak levelet, legalábbis az archívumban nem
szerepel erre utaló adat. Érdekes történeti tény, hogy ugyanabban a
társasági elismerésben részesültek, hat év eltéréssel. Darwin
egyébként fiatal korában meglehetősen idegenkedett a matematikától,
de később már másképpen fogalmazott: „Every new discovery is
mathematical in form, because there is no other guidance we can
have.” (Minden új felfedezés matematikai alakot ölt, mert más
iránymutatás nem áll rendelkezésünkre.)
<
8 Hét családot sorolt
fel: gombák, algák, mohák, páfrányok, pázsitfüvek, pálmák és
növények (plantas!) – ami mai szemmel egyáltalán nem tűnik logikus
felosztásnak.
<
9 Bár nem egyöntetű
formában, mert a növények családjainak esetében (kivételeket
megengedve) a -ceae, míg az állatoknál az -idae végződést írták elő
a nevezéktani szabályzat kidolgozói.
<
10 Más kérdés, hogy ma
már egészen eltérő elképzeléseink vannak az emlősök evolúciójáról.
<
11 Egyetlen kivétel a
Moneres, amely a gyökeret (tehát a közös őst) és az egyik ágat is
alkotja.
<
12 Hasadás során a
kiinduló populáció két új fajt eredményez, a lefűződésnél az
anyapopuláció megmarad, és a levált csoport alakul új fajjá, míg az
anagenezis esetében az anyapopuláció lassú változások sorozatával,
kihalás nélkül megy át egy másikba.
<
13 „Az élet fáját talán
az élet koralljának kellene nevezni, melynek alsó ágai már nem
élnek, így nincsenek közöttük átkötések.”
<
14 Első komoly, pár
évvel a fentiek után megjelent monográfiájának éppen ez volt a
témája: The Structure and Distribution of Coral Reefs… (Darwin,
1842). Az általa név szerint említett korallok jelentős része
(például Porites és Seriatopora) egyébként anasztomizálhat is!
<
|
|